Deep analysis for high-volatility longevity claims.
Extended essays that separate validated signal from narrative acceleration across longevity, AI biology, and clinical translation.
Explore Longform Pillars
These pillar pages group the current catalog by durable topic so readers and crawlers can move from one article into a full subfield.
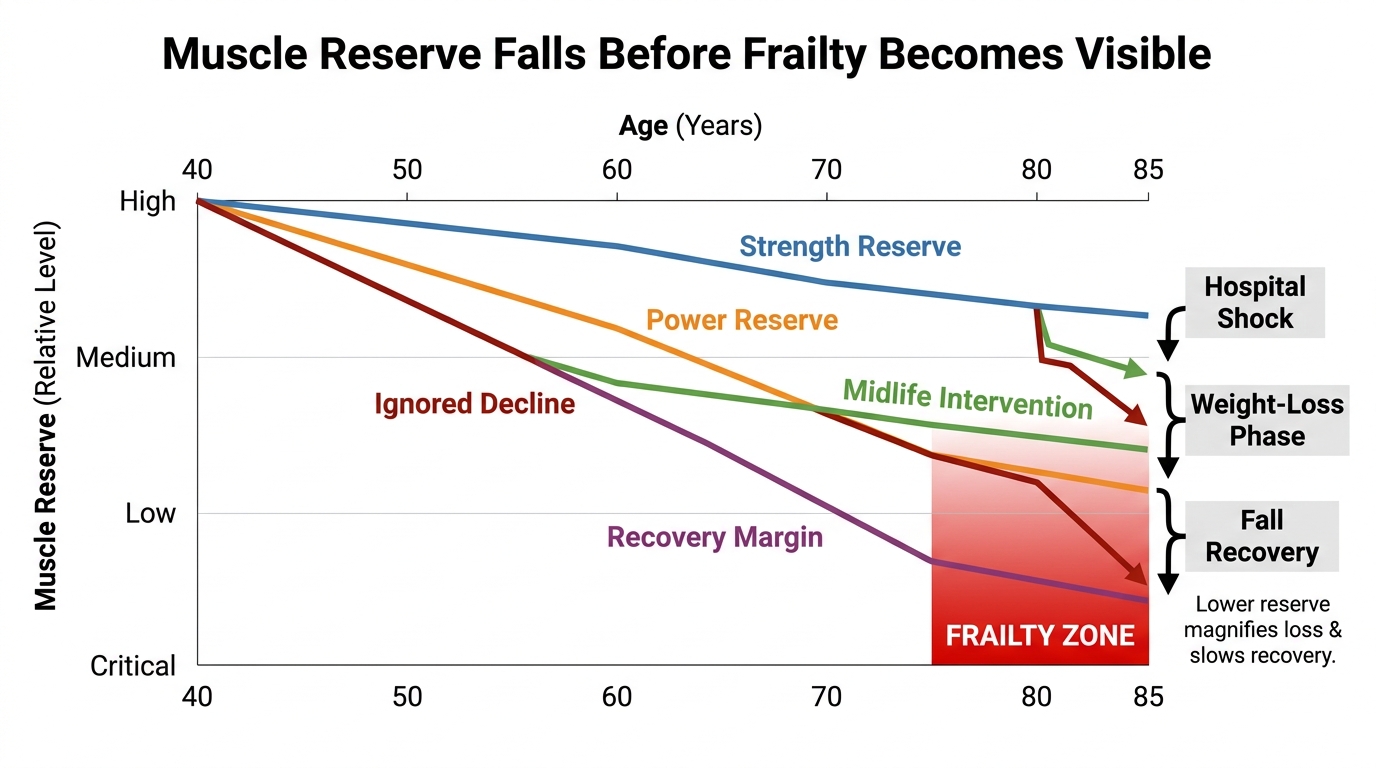
Sarcopenia Before Frailty: The Real Midlife Longevity Pivot
Why muscle decline becomes a longevity problem long before visible frailty, why strength and power matter more than aesthetics, and why midlife is the decisive prevention window.

Measuring ROI on Longevity Interventions
A 2026 LifeMeter analysis of how to measure return on longevity interventions without confusing biomarker movement, marketing confidence, and real gains in healthy life.
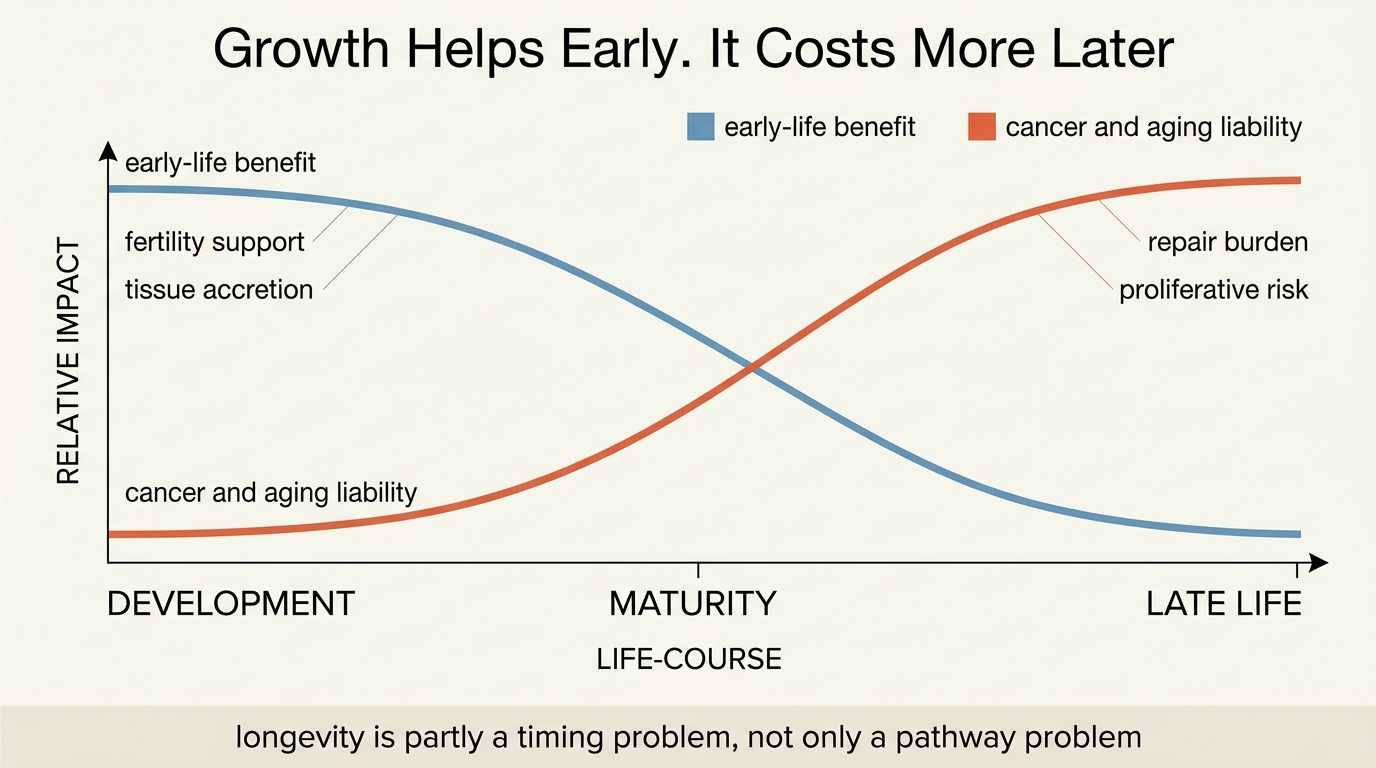
Tradeoffs in Longevity: Cancer Risk, Fertility, and Growth
A 2026 LifeMeter analysis of why some of the same growth programs that help organisms build, reproduce, and repair early in life can become liabilities for cancer risk and longevity later on.
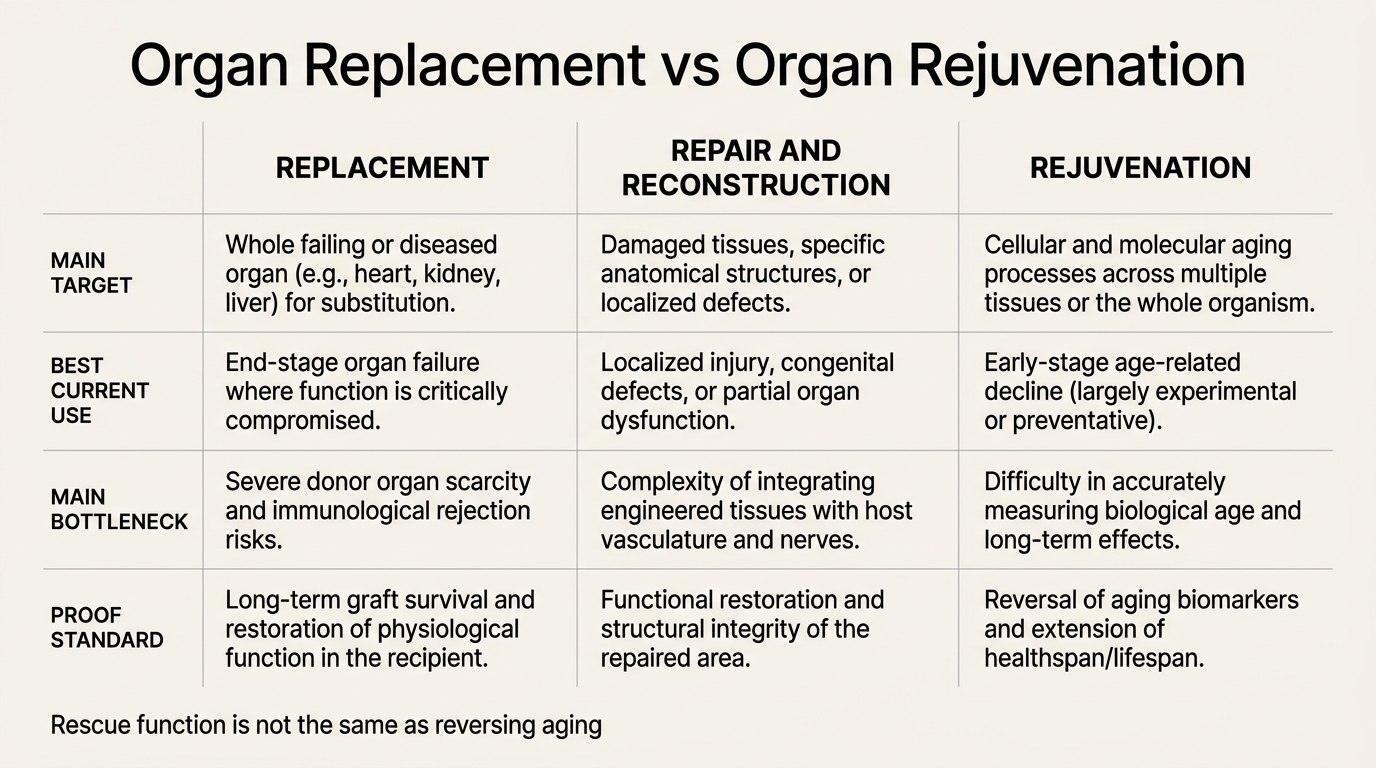
Organ Replacement vs Organ Rejuvenation
A 2026 LifeMeter analysis of where organ replacement already works, where organ rejuvenation still faces older-host biology, and why those are not interchangeable longevity strategies.
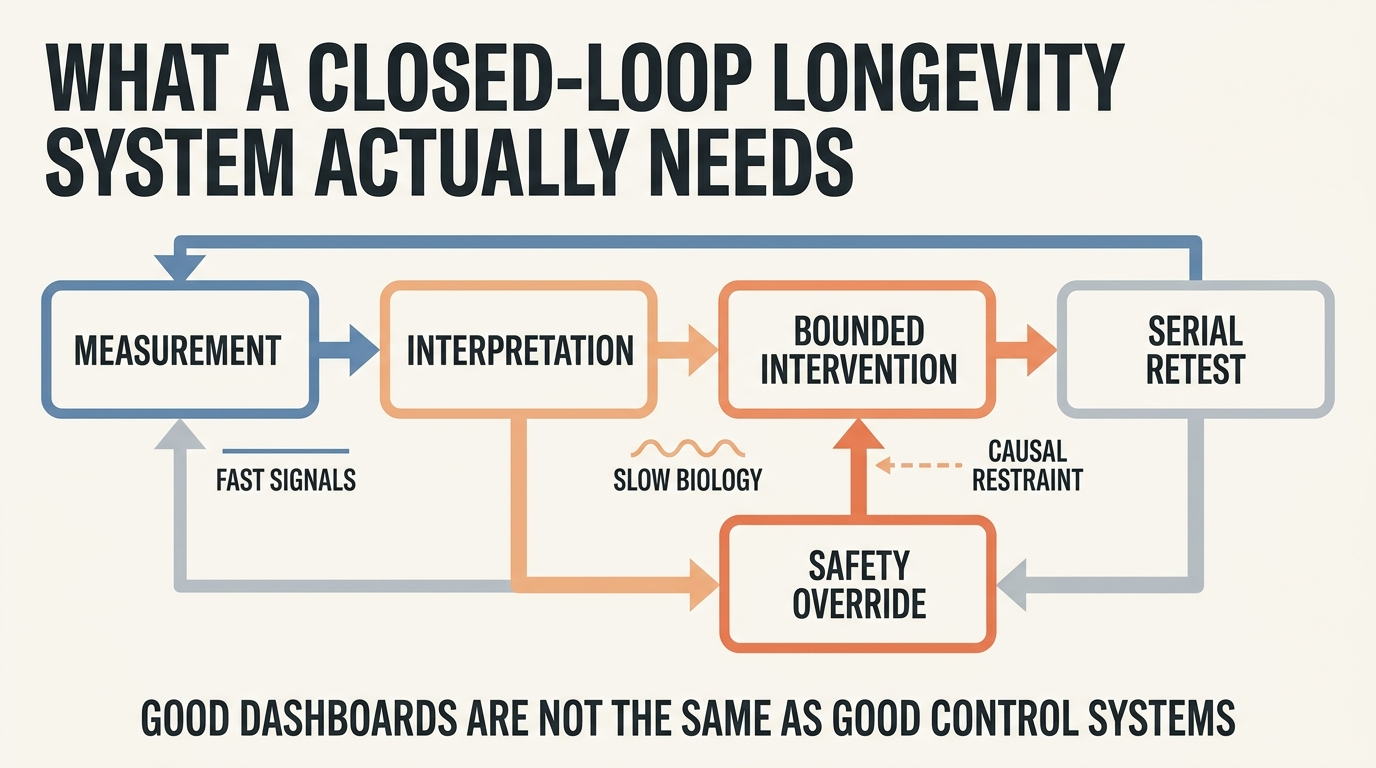
Closed-Loop Longevity Systems: Measure, Intervene, Adapt
A 2026 LifeMeter analysis of what a real closed-loop longevity system would require, where the model already works, and why most anti-aging feedback claims still outrun measurement quality.
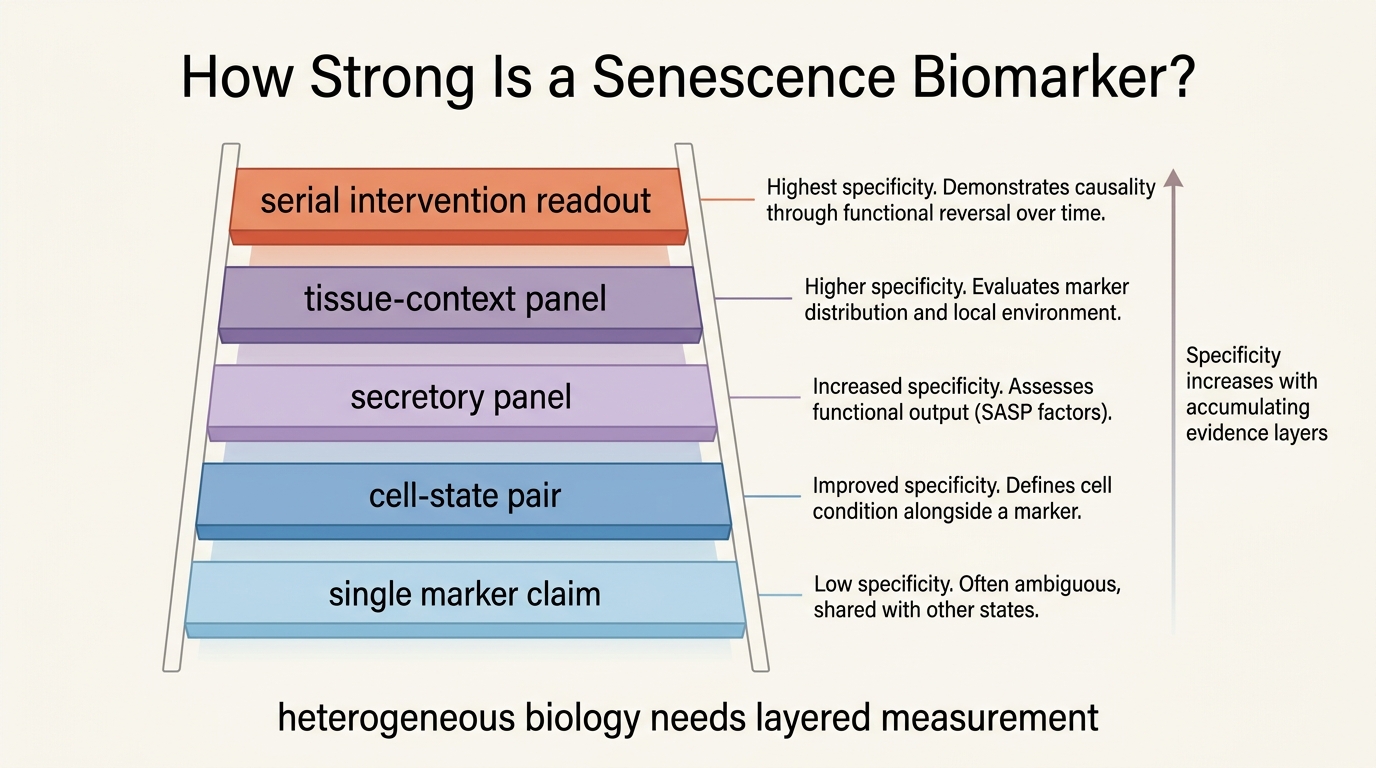
Biomarkers for Senescent Cell Burden
A 2026 LifeMeter analysis of how senescent cell burden is actually measured, why no single biomarker is sufficient, and which marker combinations are closest to decision-grade use.
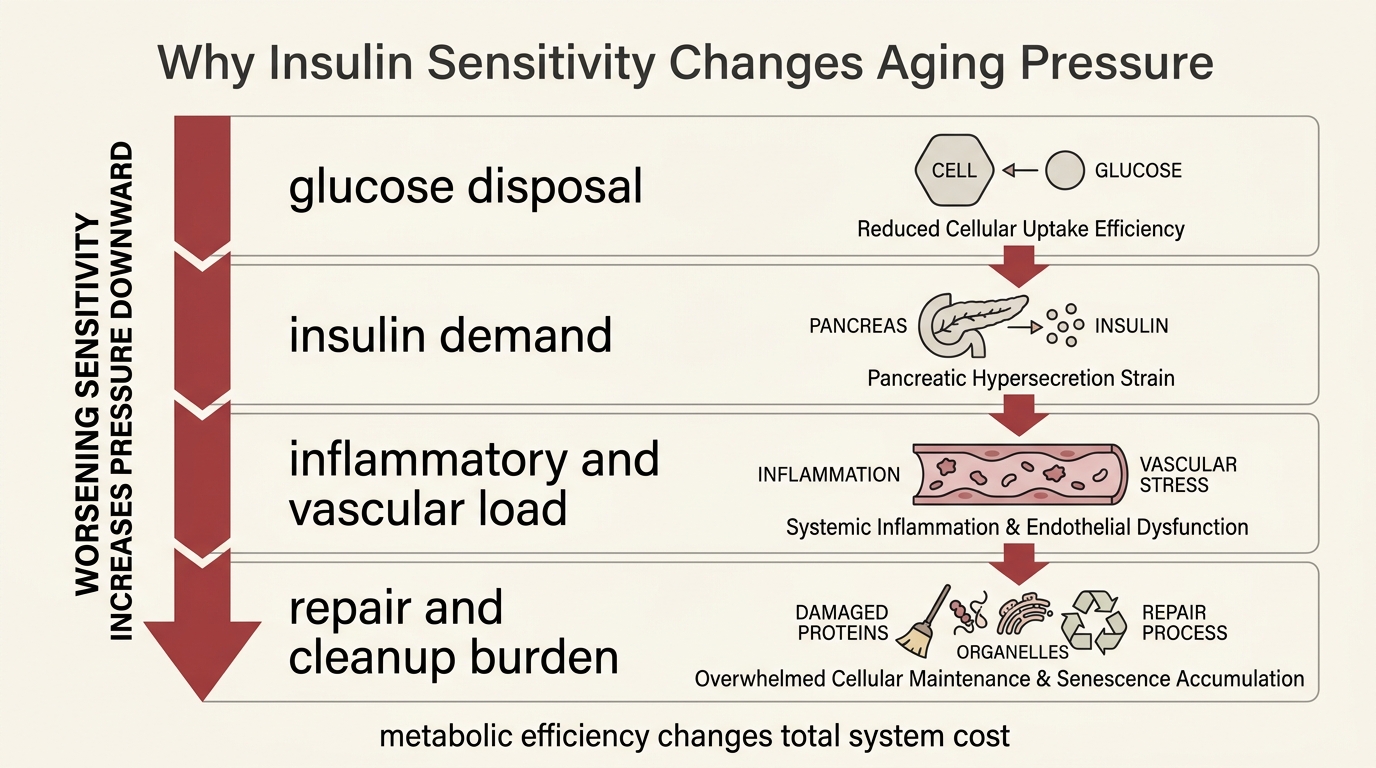
Insulin Sensitivity as a Longevity Lever
A 2026 LifeMeter analysis of why insulin sensitivity matters because it lowers metabolic stress, preserves tissue flexibility, and changes disease risk, while still falling short of a standalone human longevity proof.
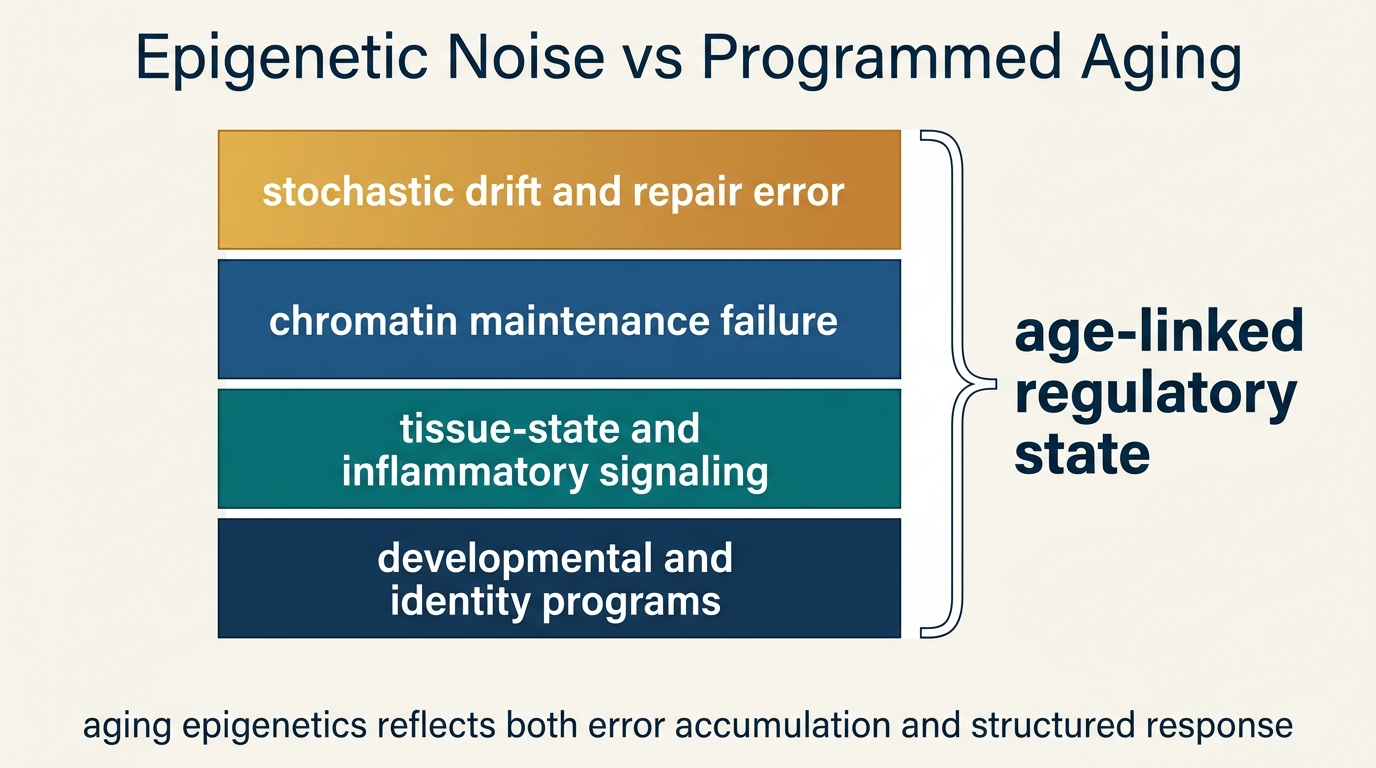
Epigenetic Noise vs Programmed Aging
A LifeMeter analysis of whether age-linked methylation drift reflects stochastic epigenetic noise, coordinated aging programs, or a layered interaction between the two.
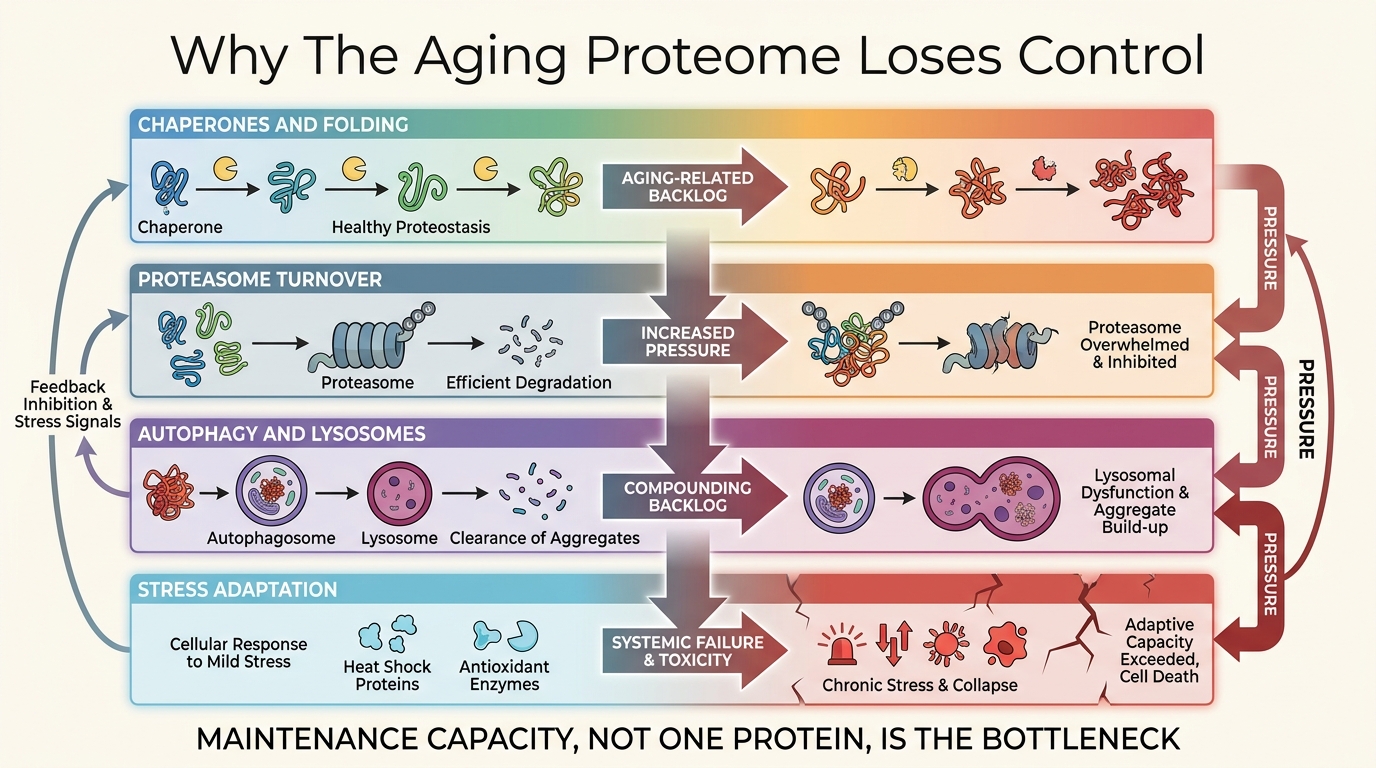
Proteostasis Collapse and the Aging Proteome
A LifeMeter analysis of why protein quality control fails with age, how that failure spreads across tissues, and why proteostasis is better treated as a maintenance layer than a single drug target.
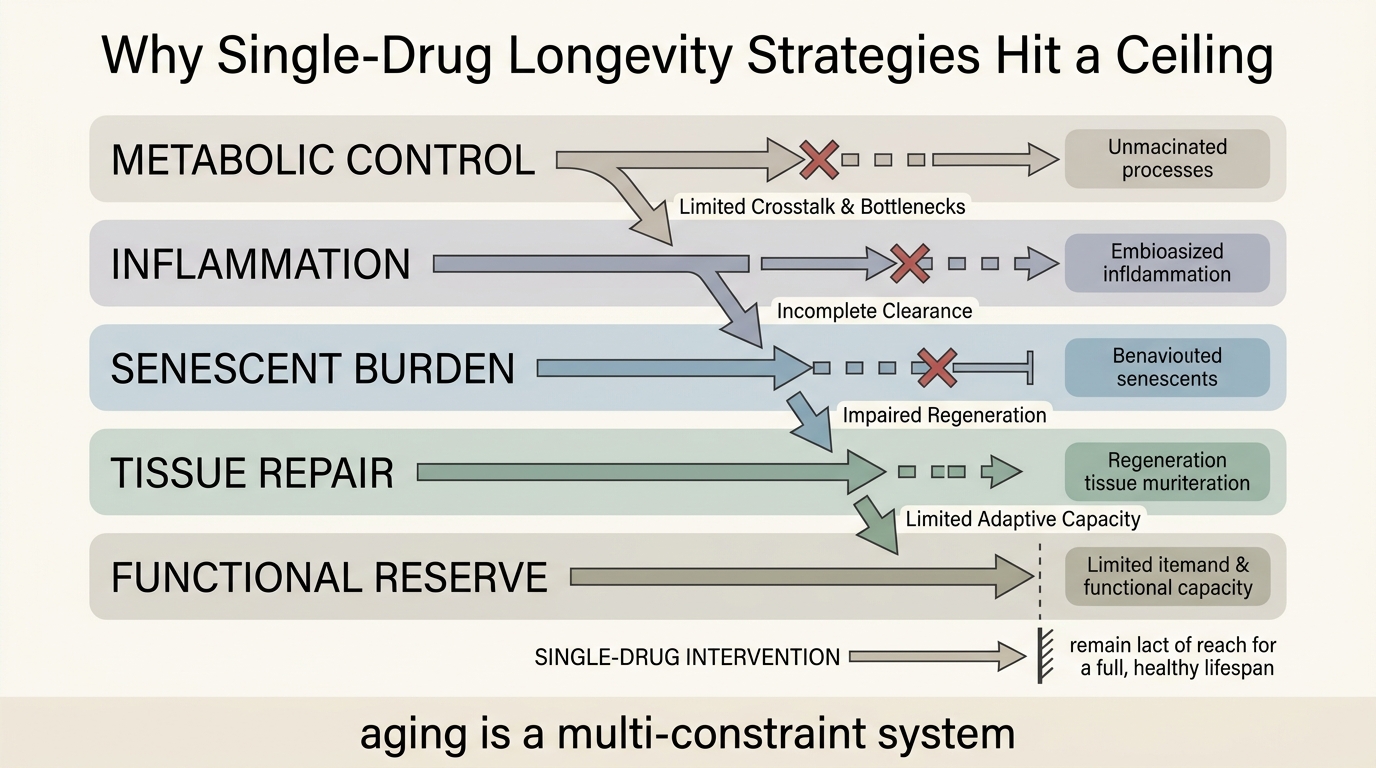
Combination Therapies: Why Single-Drug Approaches Fail
A LifeMeter analysis of why aging rarely yields to single-pathway intervention, where combination therapy logic is strongest, and what makes multi-agent longevity treatment hard to validate and govern.
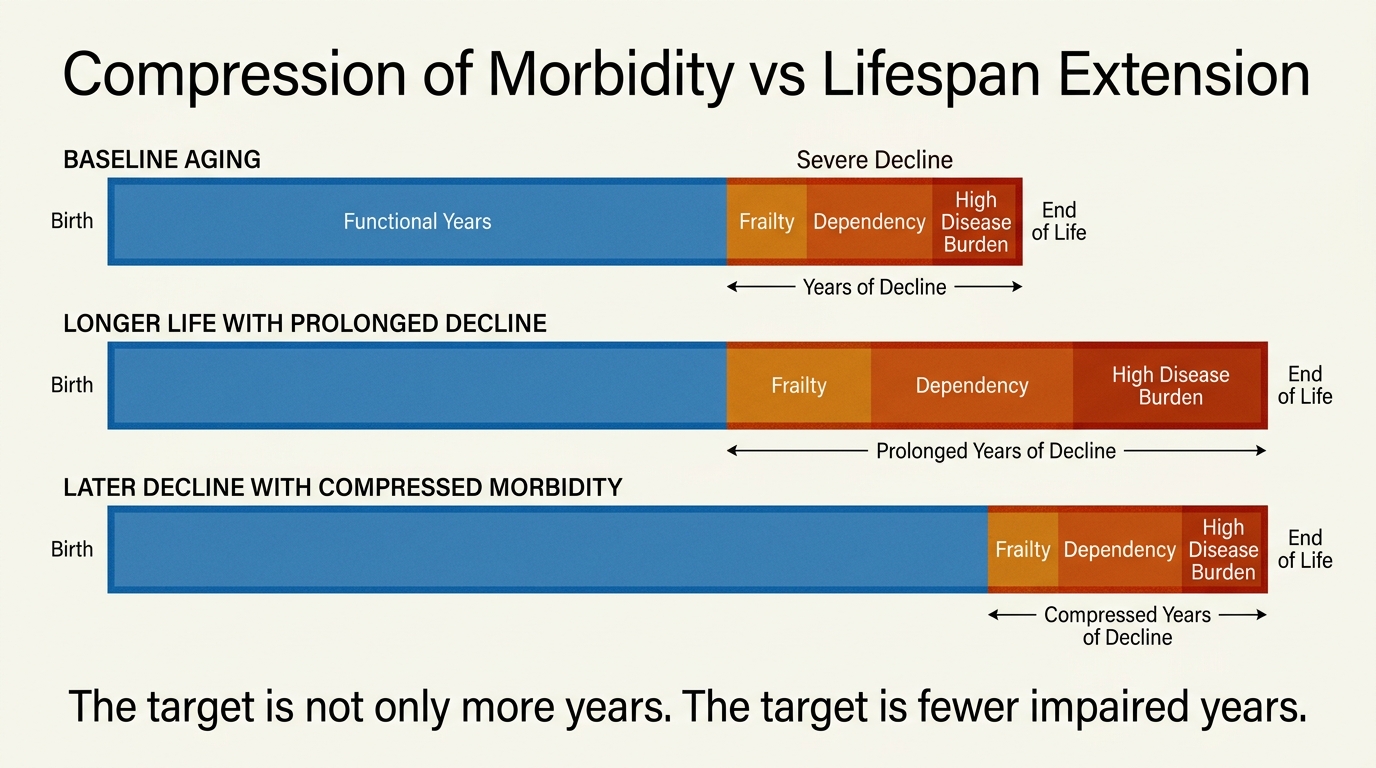
Compression of Morbidity vs Lifespan Extension
A LifeMeter analysis of the difference between extending lifespan and compressing morbidity, with a practical framework for judging whether a longevity intervention improves years alive, years functional, or both.
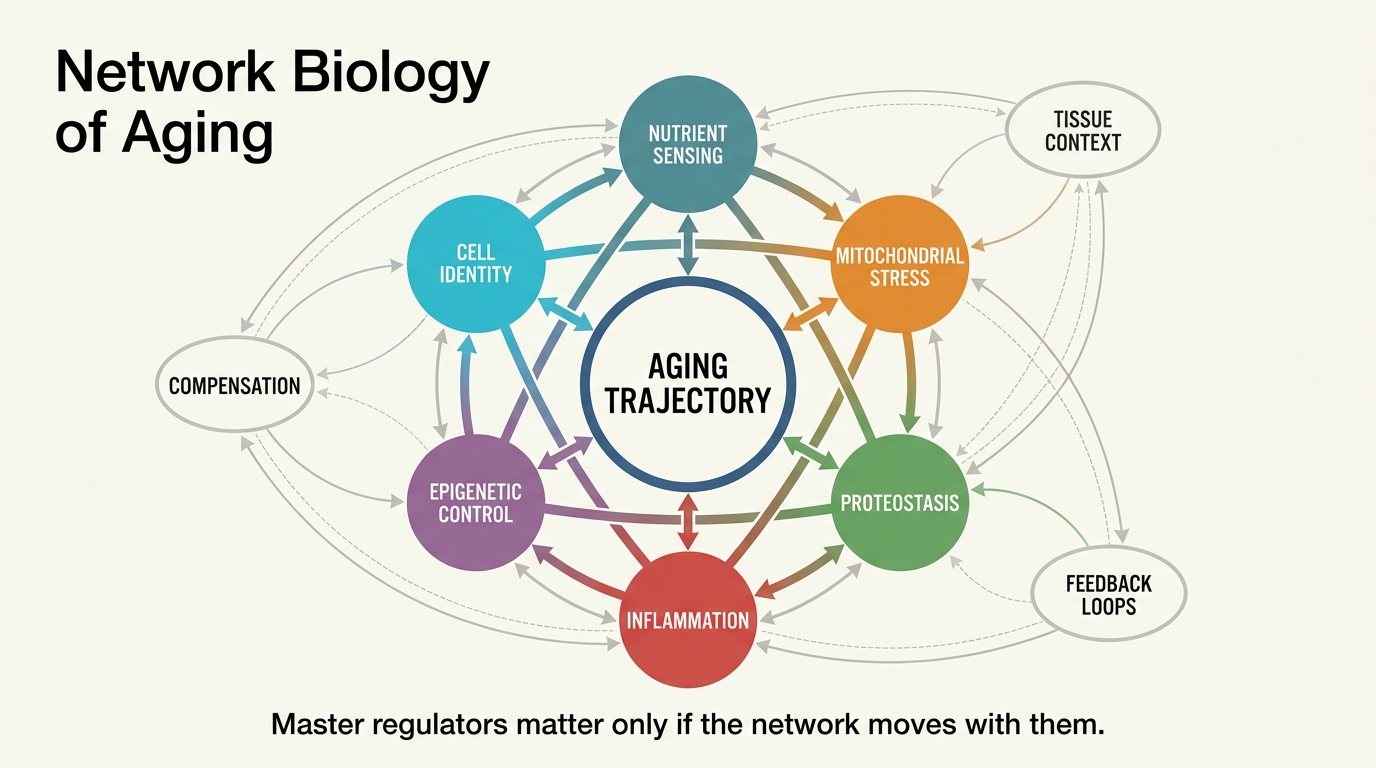
Network Biology of Aging: Identifying Master Regulators
A LifeMeter analysis of network biology of aging, explaining what master regulators can reveal, where systems maps are strongest, and why one-node rejuvenation stories usually outrun the evidence.
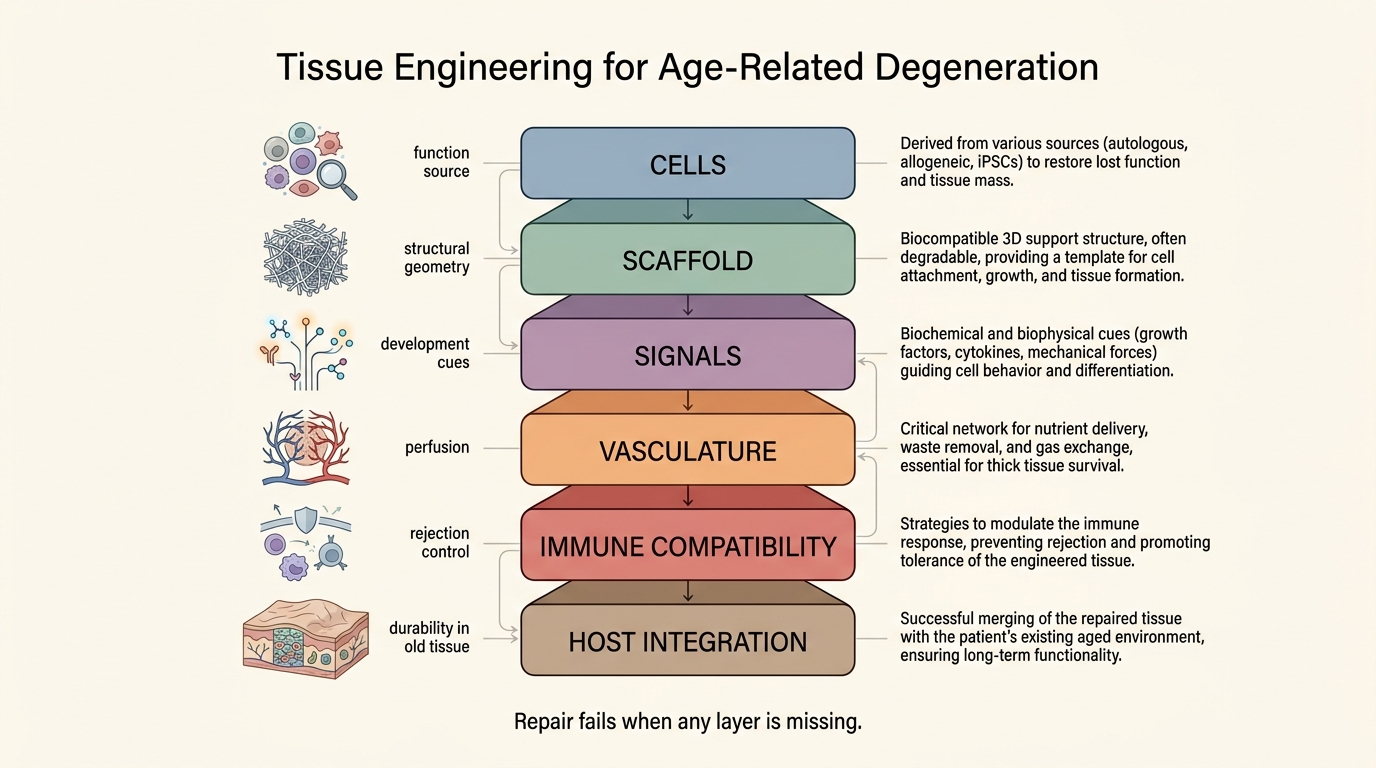
Tissue Engineering for Age-Related Degeneration
A 2026 LifeMeter analysis of tissue engineering for age-related degeneration, separating real repair progress from the much harder problem of rebuilding old organs inside old biological environments.
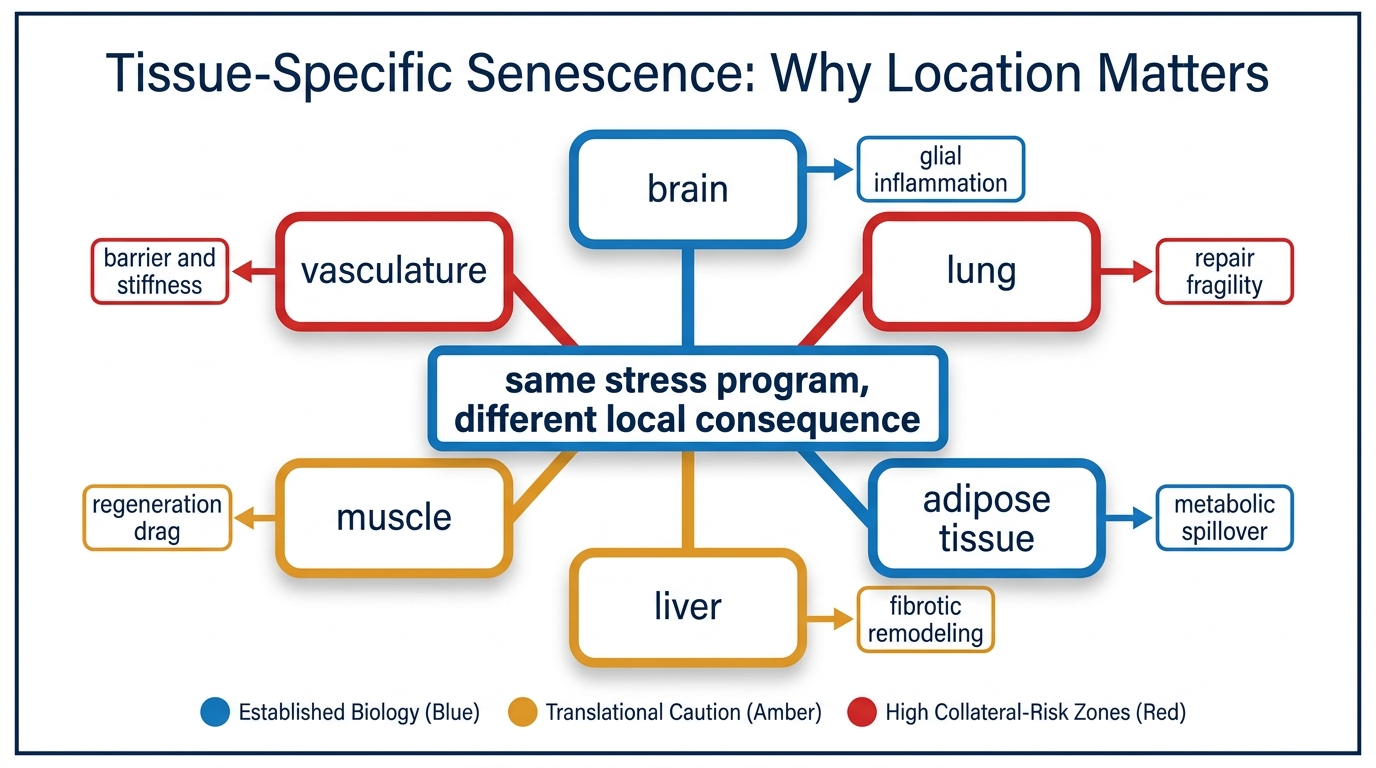
Tissue-Specific Senescence: Why Location Matters
A 2026 LifeMeter analysis of why senescence is not one uniform burden, how tissue location changes biological meaning, and why intervention logic has to follow local context rather than a generic clearance story.
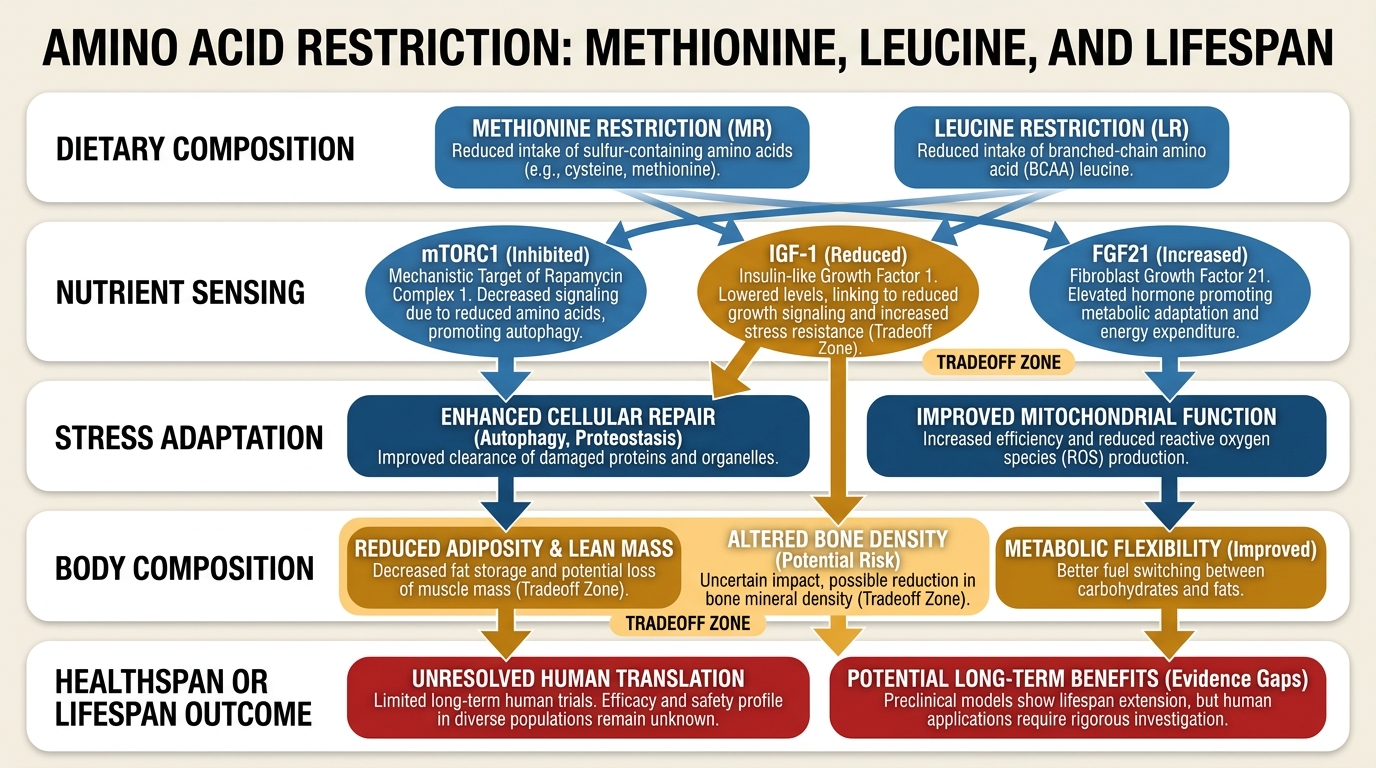
Amino Acid Restriction: Methionine, Leucine, and Lifespan
A LifeMeter analysis of what methionine and leucine restriction actually show in aging biology, where the animal signal is strongest, and why human translation remains narrow and conditional.
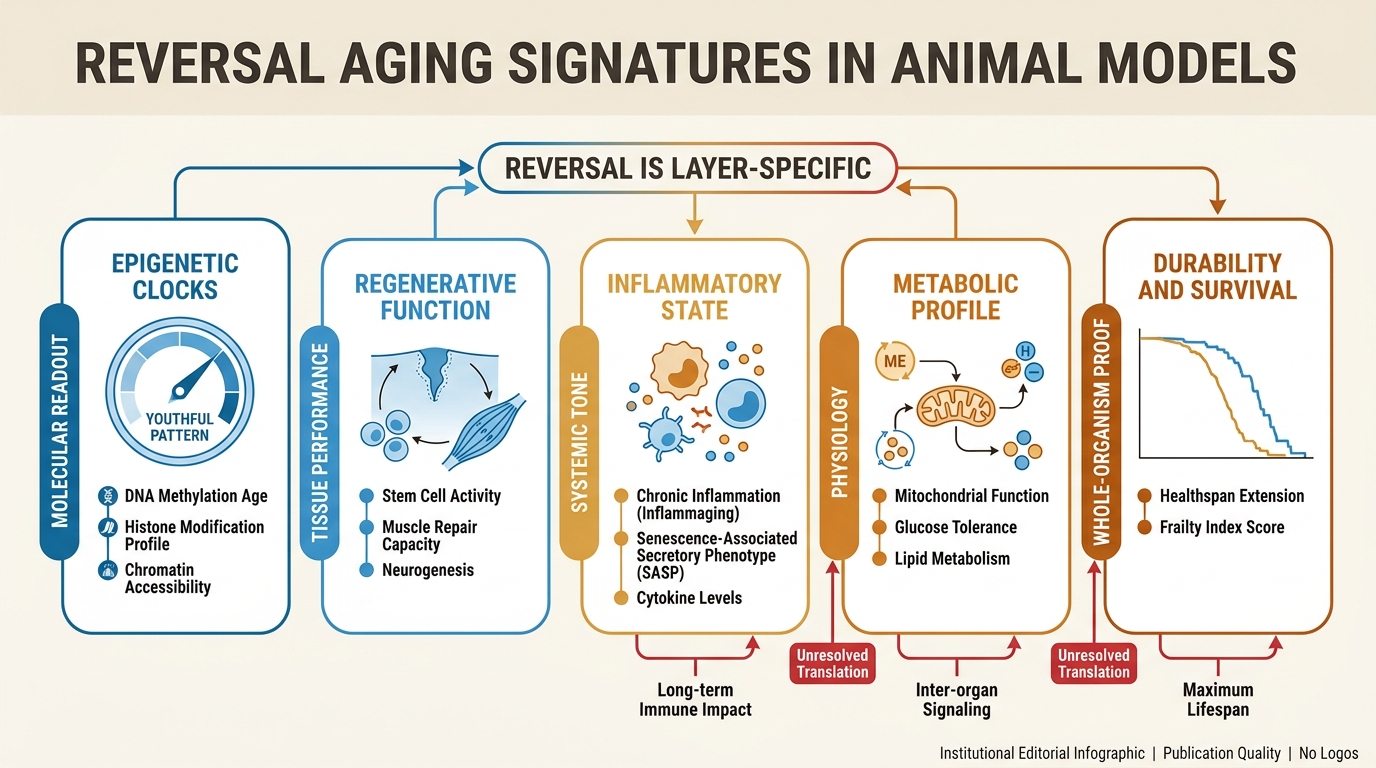
Reversible Aging Signatures in Animal Models
A LifeMeter analysis of what animal models actually show when age-linked traits move backward, which signals look reversible first, and why that still falls short of a general age-reversal verdict.
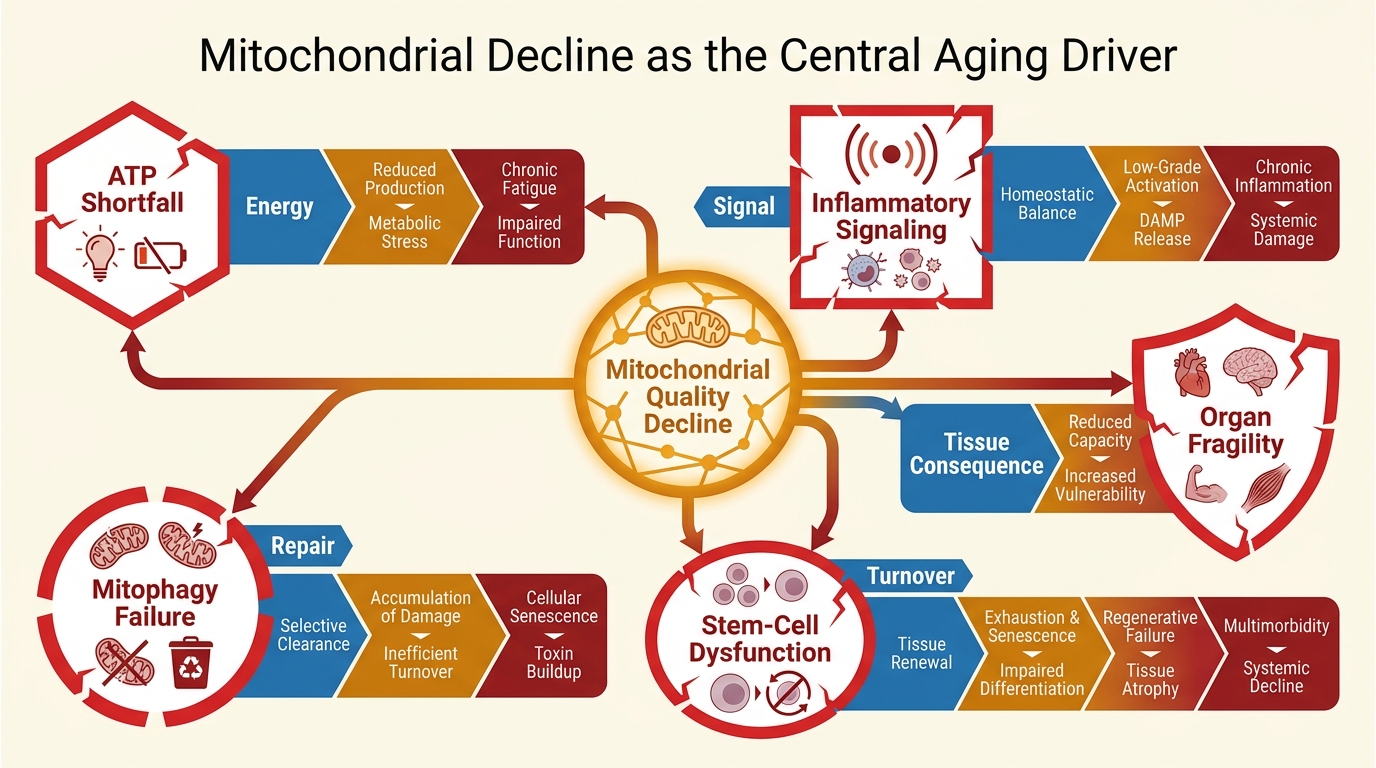
Mitochondrial Decline as the Central Aging Driver
A LifeMeter analysis of why mitochondrial decline matters across aging, where the central-driver claim is justified, and where it overreaches the evidence.
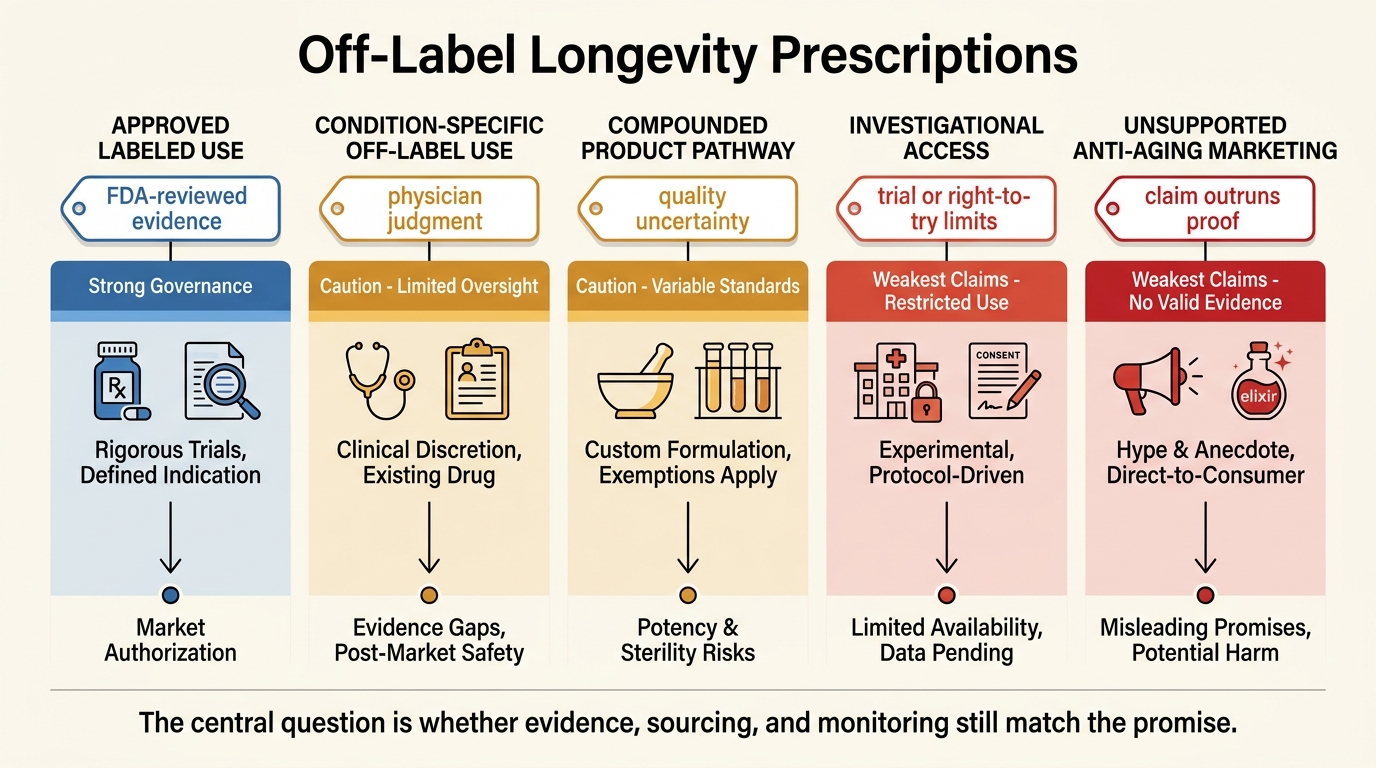
Off-Label Longevity Prescriptions: Risks and Patterns
A LifeMeter analysis of off-label longevity prescribing, separating ordinary medical discretion from the weaker evidence, compounding risk, and governance gaps common in the anti-aging market.
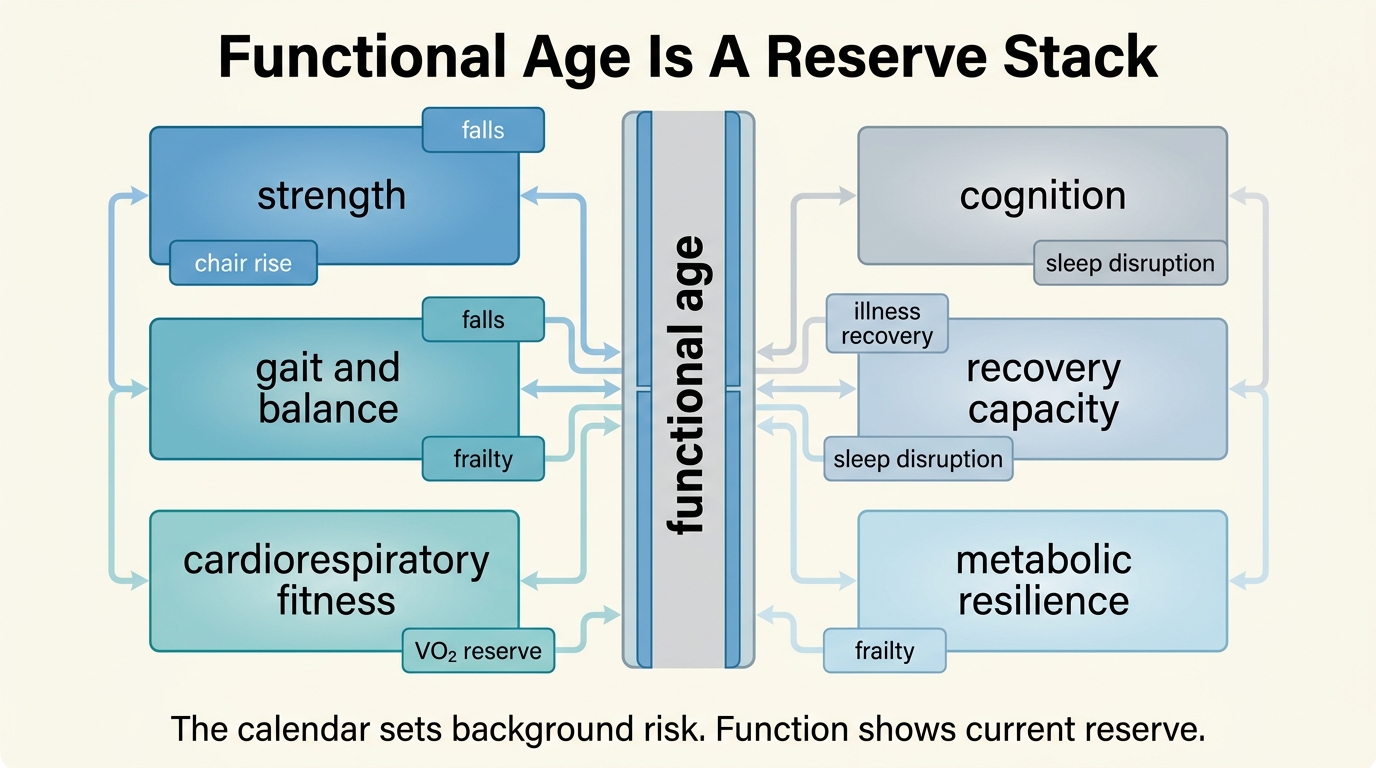
Functional Age vs Chronological Age: Which Matters More?
A LifeMeter analysis of why functional age often predicts real-world decline better than the calendar alone, where biological clocks help, and why the right model is layered rather than competitive.
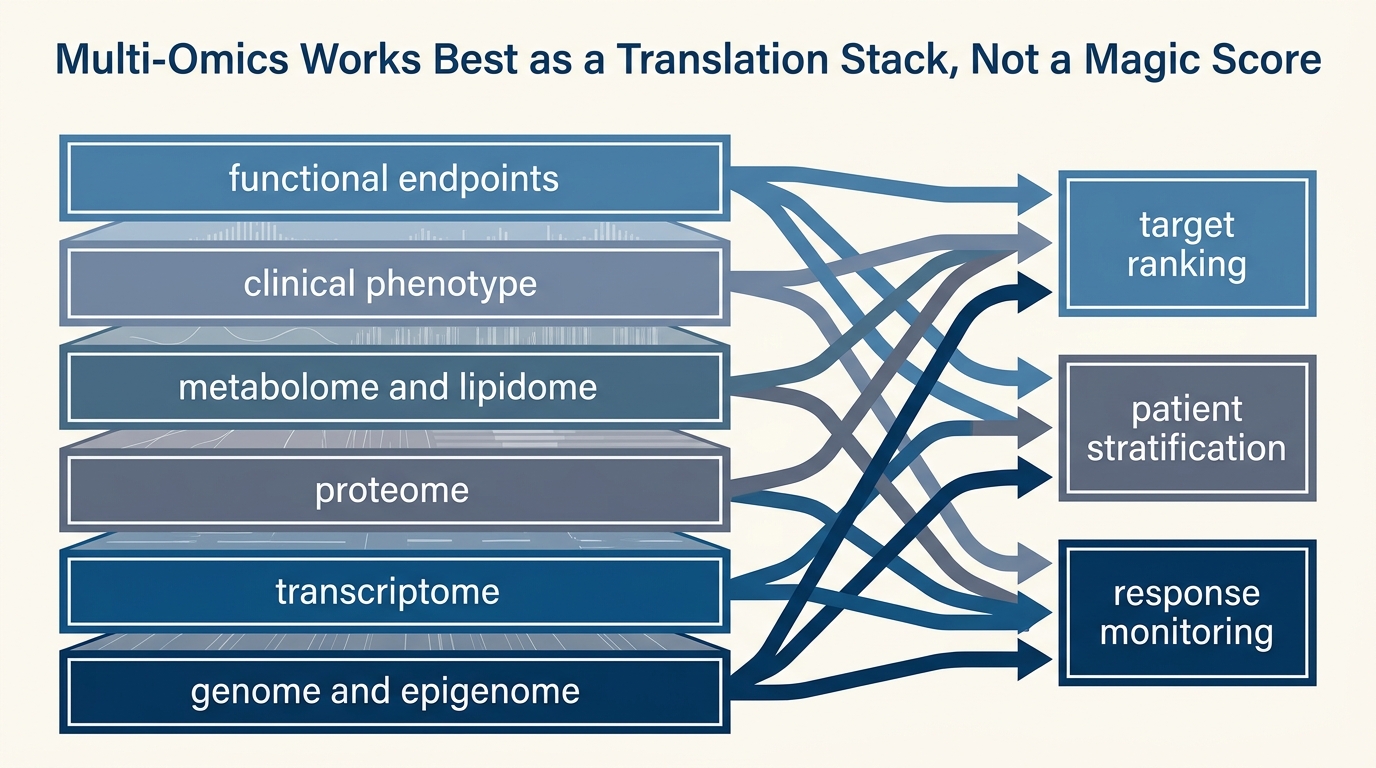
Multi-Omics Integration: From Data to Intervention
A LifeMeter analysis of what multi-omics integration can already do in aging research, where it is strongest for stratification and monitoring, and why turning signal into intervention still requires a narrower evidentiary bridge.
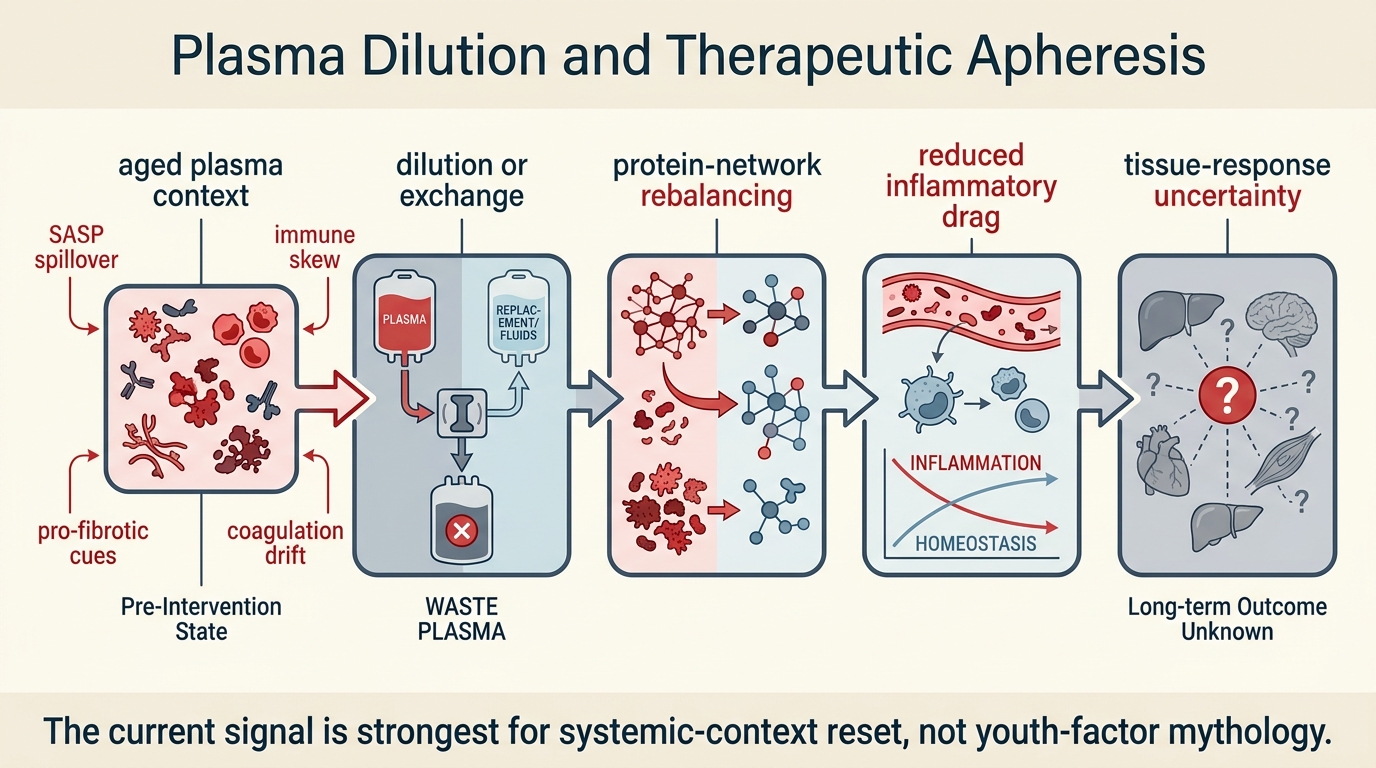
Plasma Dilution and Therapeutic Apheresis
A LifeMeter analysis of plasma dilution and therapeutic apheresis, separating the real mechanistic signal around circulating-factor reset from the still-limited human evidence for broad rejuvenation.
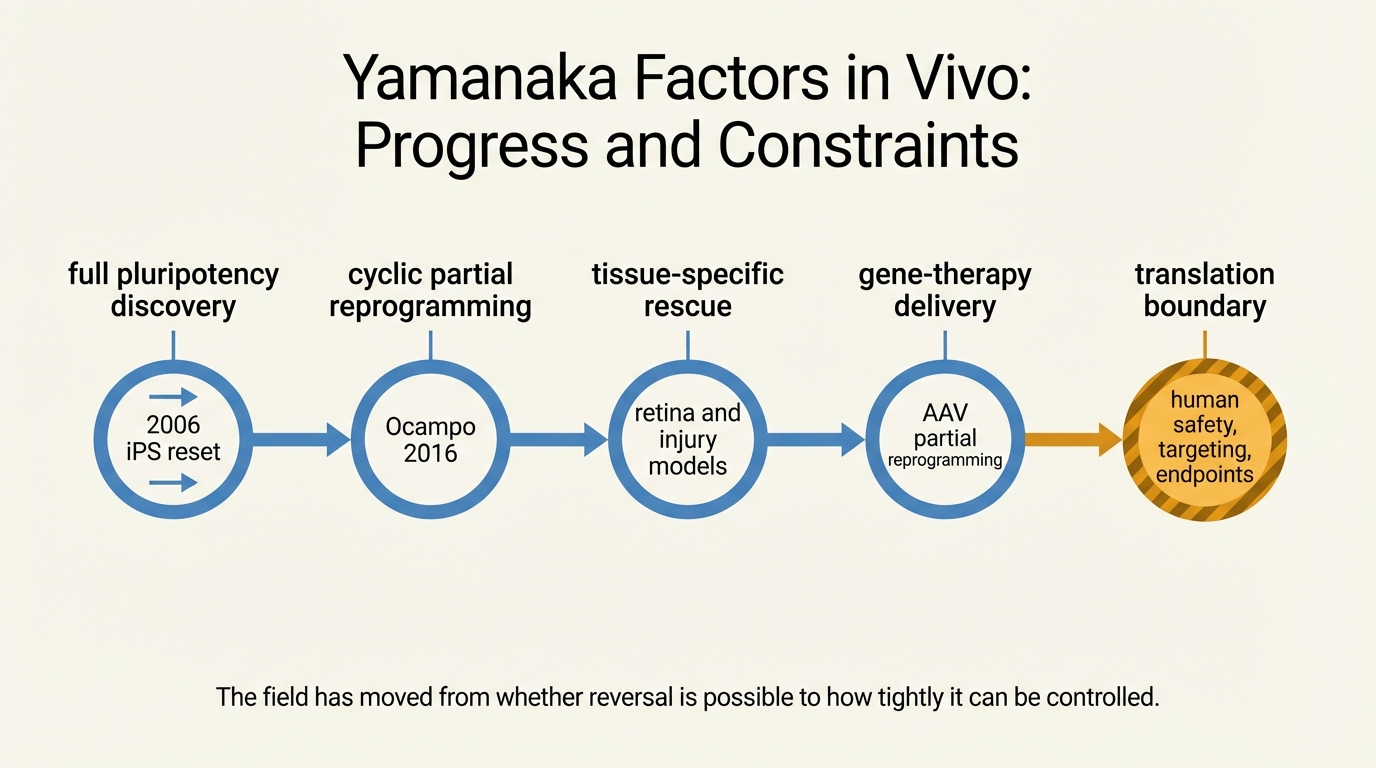
Yamanaka Factors in Vivo: Progress and Constraints
A 2026 LifeMeter audit of in vivo Yamanaka-factor research, separating the real regenerative signal from the still-unresolved control, surveillance, and endpoint constraints.
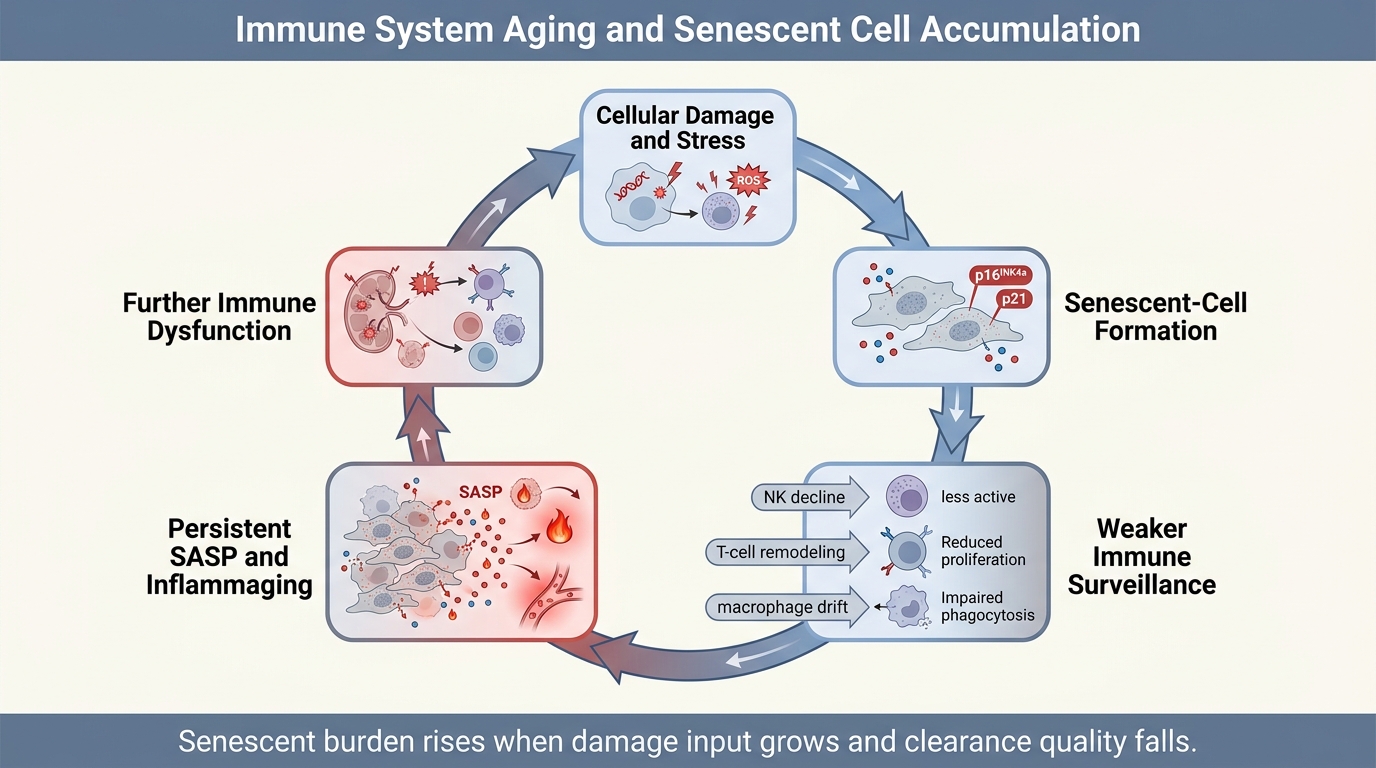
Immune System Aging and Senescent Cell Accumulation
A 2026 LifeMeter analysis of how immune aging weakens senescent-cell clearance, why that failure accelerates inflammaging, and which parts of the senescence-control story are established versus still unresolved.
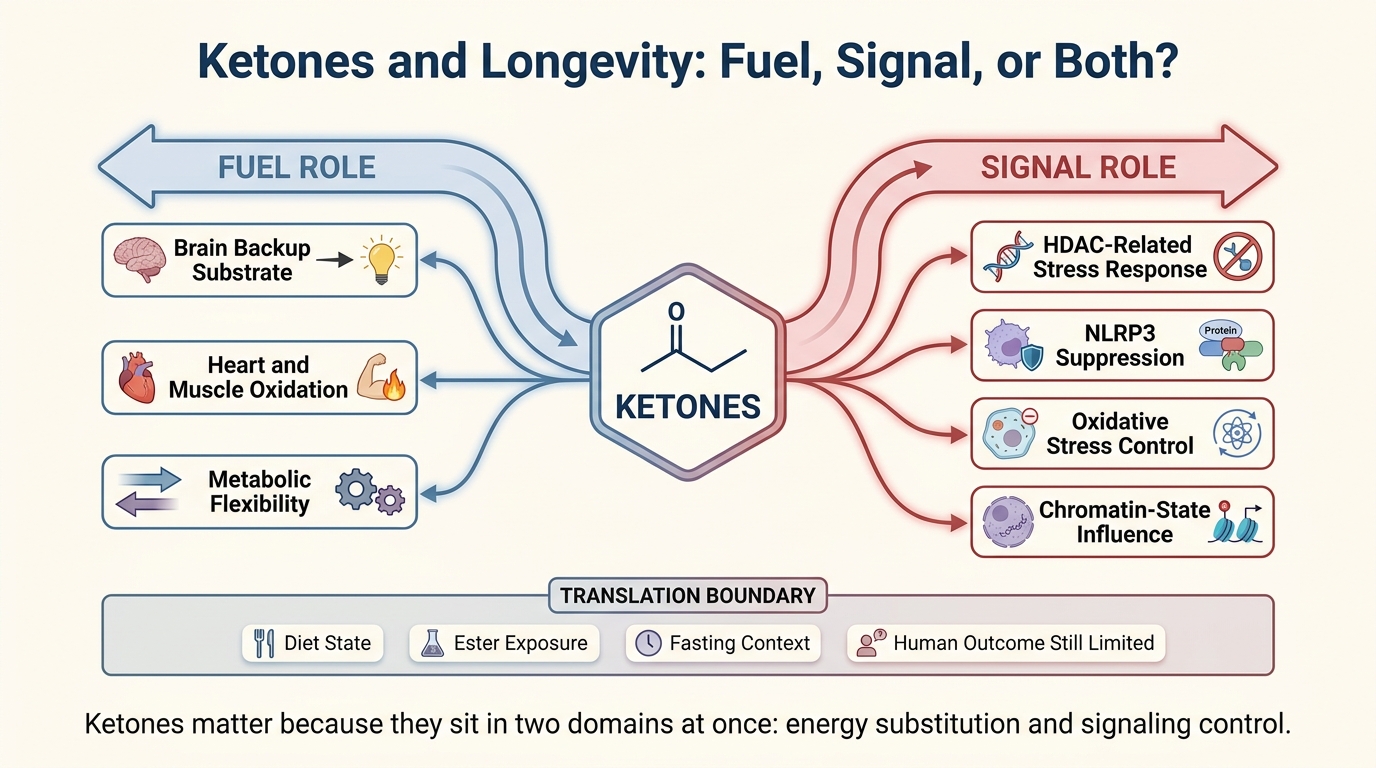
Ketones and Longevity: Fuel, Signal, or Both?
A 2026 LifeMeter analysis of ketones in aging biology, separating their real role as fuel and signaling metabolites from the much stronger claim that ketosis already delivers broad human longevity gains.
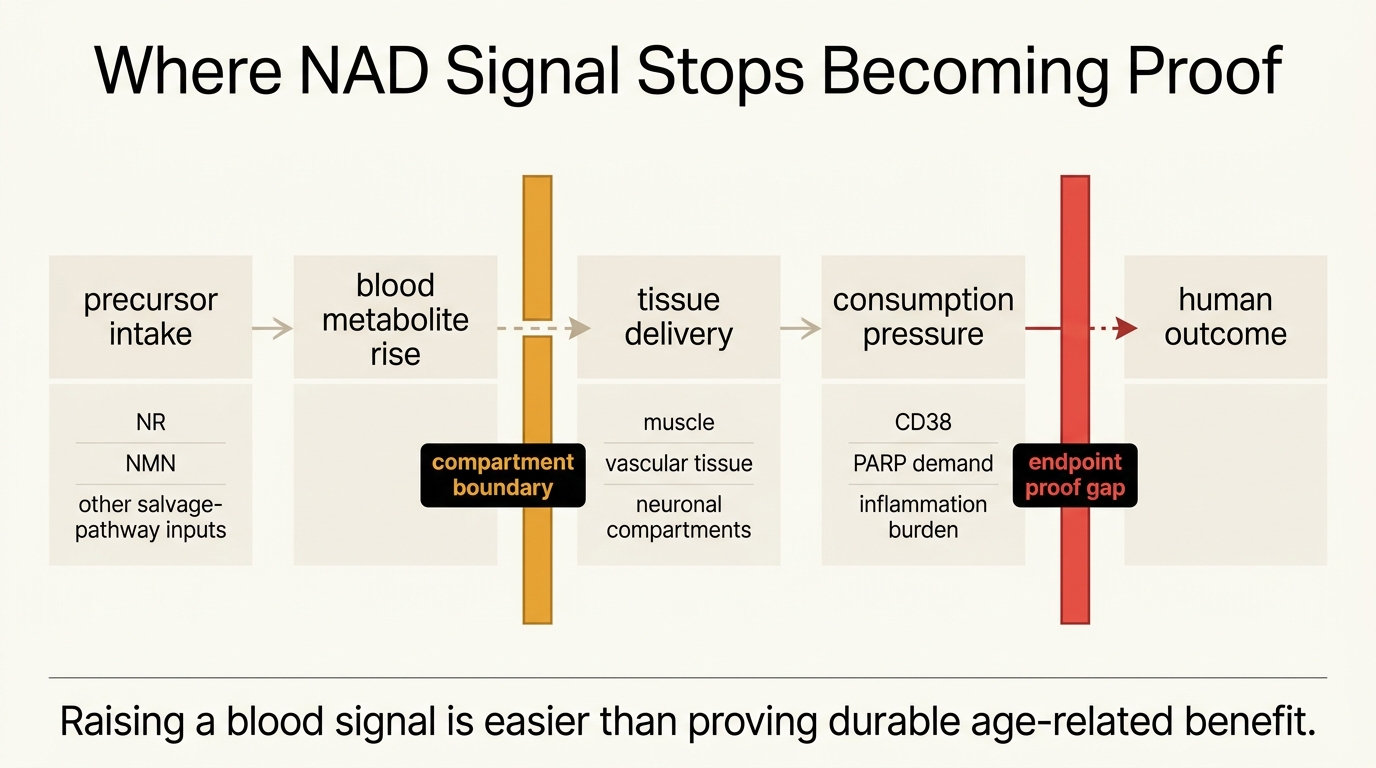
NAD+ Restoration: Mechanism, Hype, and Clinical Signal
A 2026 LifeMeter audit of NAD+ restoration claims, separating the strong mechanistic case for NAD biology from the still-limited human evidence for broad anti-aging benefit.
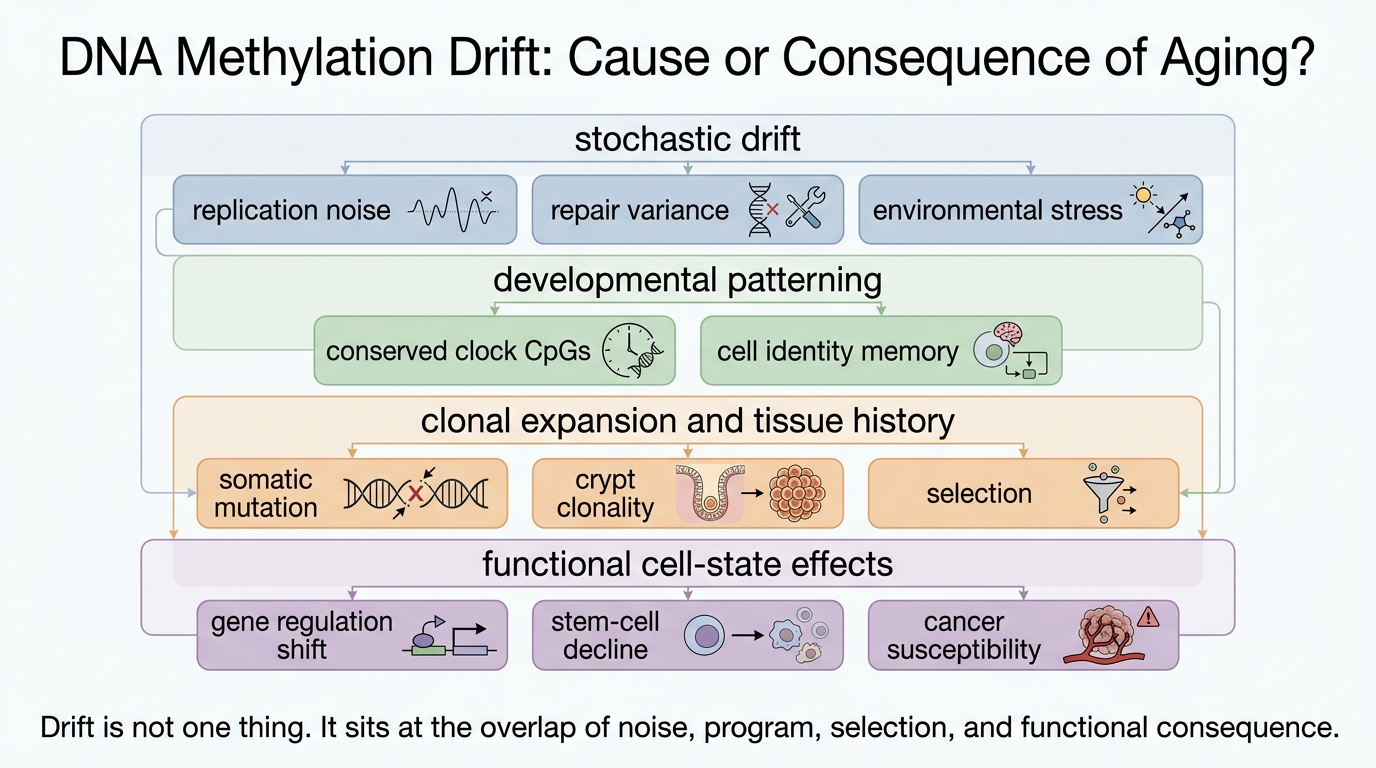
DNA Methylation Drift: Cause or Consequence of Aging?
A 2026 LifeMeter analysis of whether age-linked DNA methylation drift merely records biological wear or also helps drive aging outcomes through cell identity loss, clonal expansion, and repair failure.
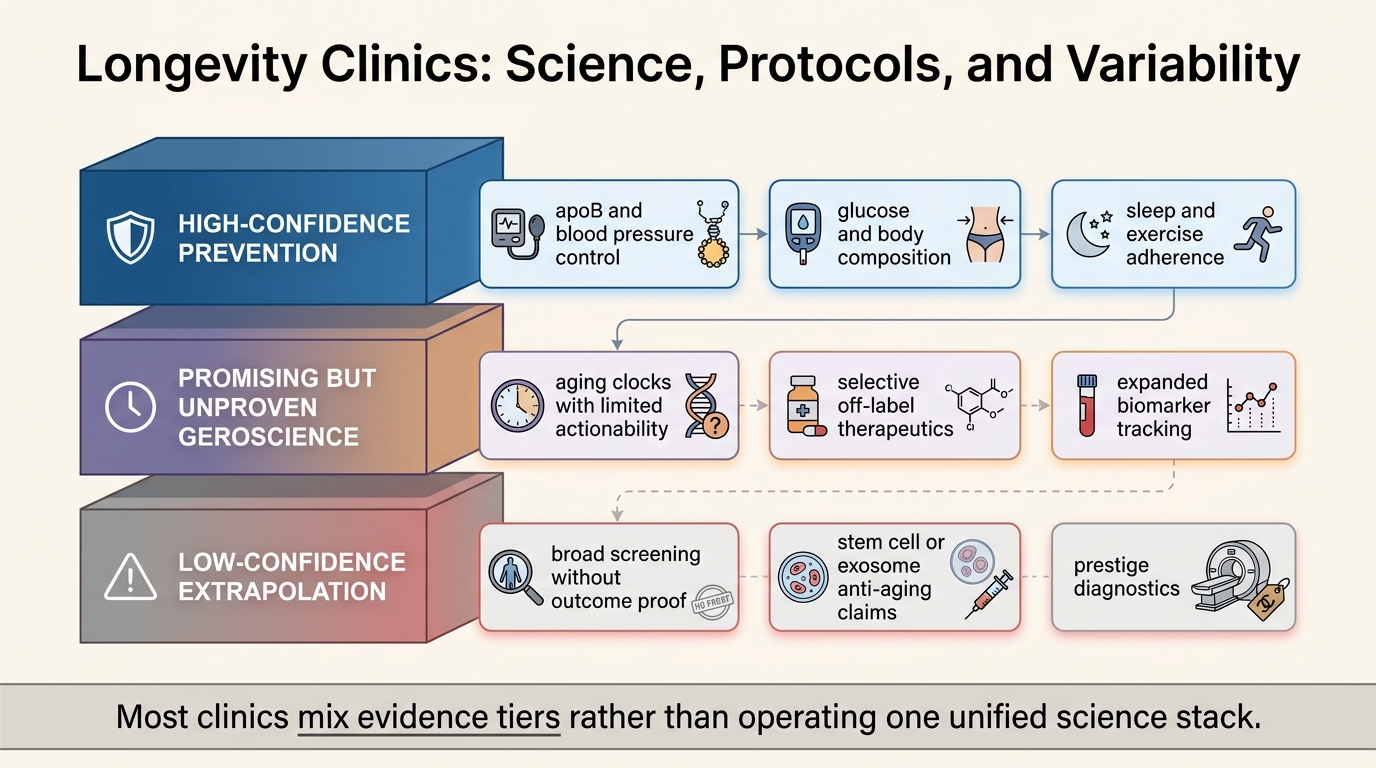
Longevity Clinics: Science, Protocols, and Variability
A 2026 LifeMeter refresh on what longevity clinics actually deliver, which parts rest on established preventive medicine, and where protocol variability reflects weaker aging-specific evidence.

Sirtuins in Humans: From Yeast Biology to Clinical Relevance
Sirtuins became central to longevity biology because the underlying enzymes are real. The unresolved question is how much of the early lifespan signal actually survives translation into humans.
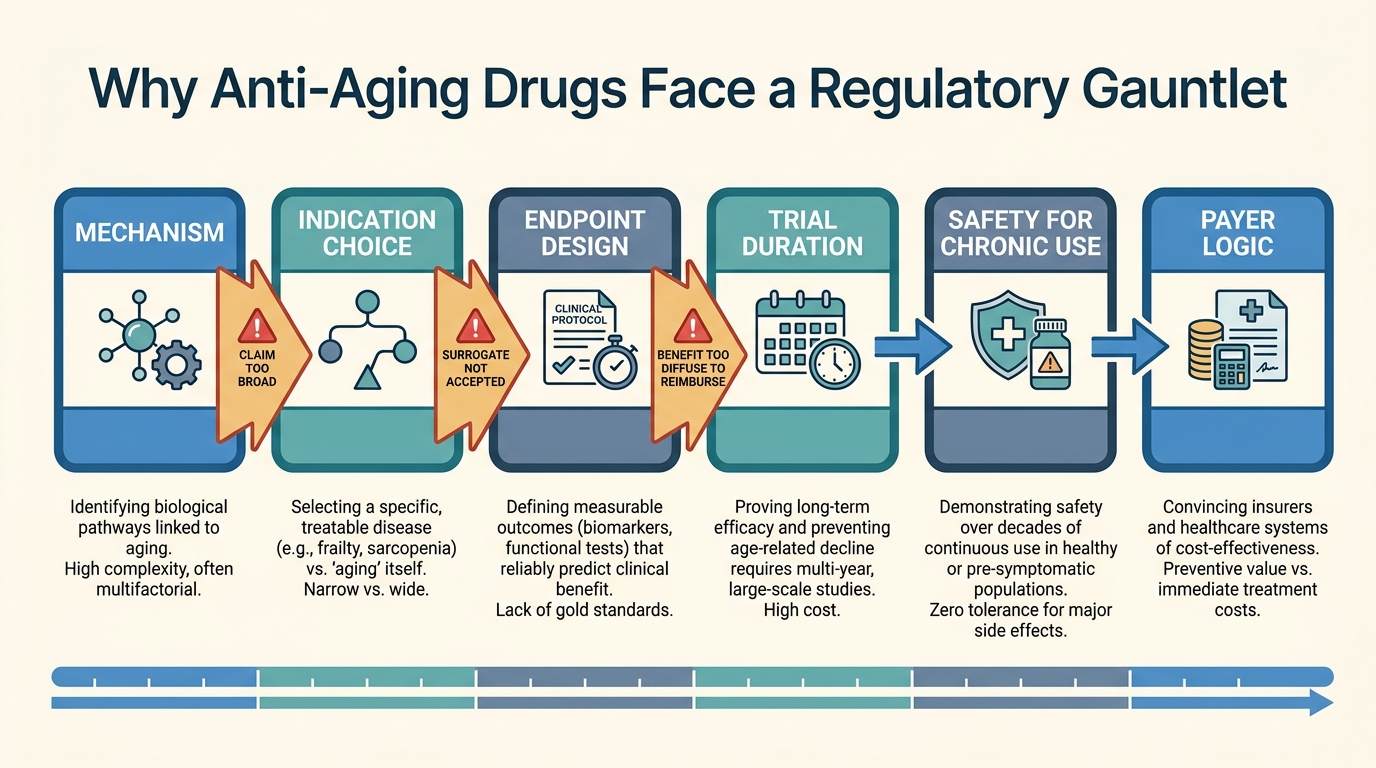
Regulatory Barriers to Anti-Aging Drugs
Why anti-aging drug programs still face an unusually hard path through indication design, endpoint validation, and payer logic even when the biology looks plausible.
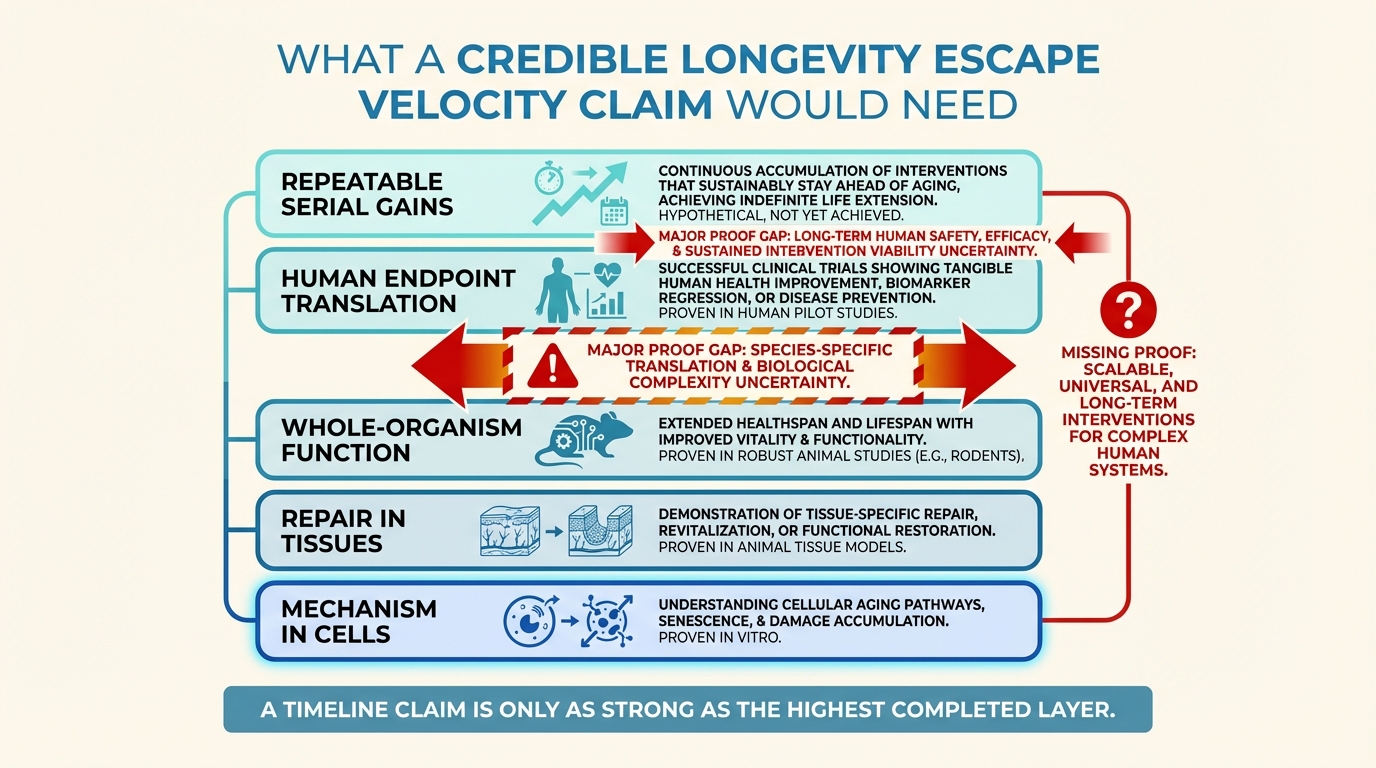
Longevity Escape Velocity: Realistic or Misleading?
A disciplined read of longevity escape velocity as a strategic concept, where it remains useful, and why it becomes misleading once timelines outrun current repair evidence.
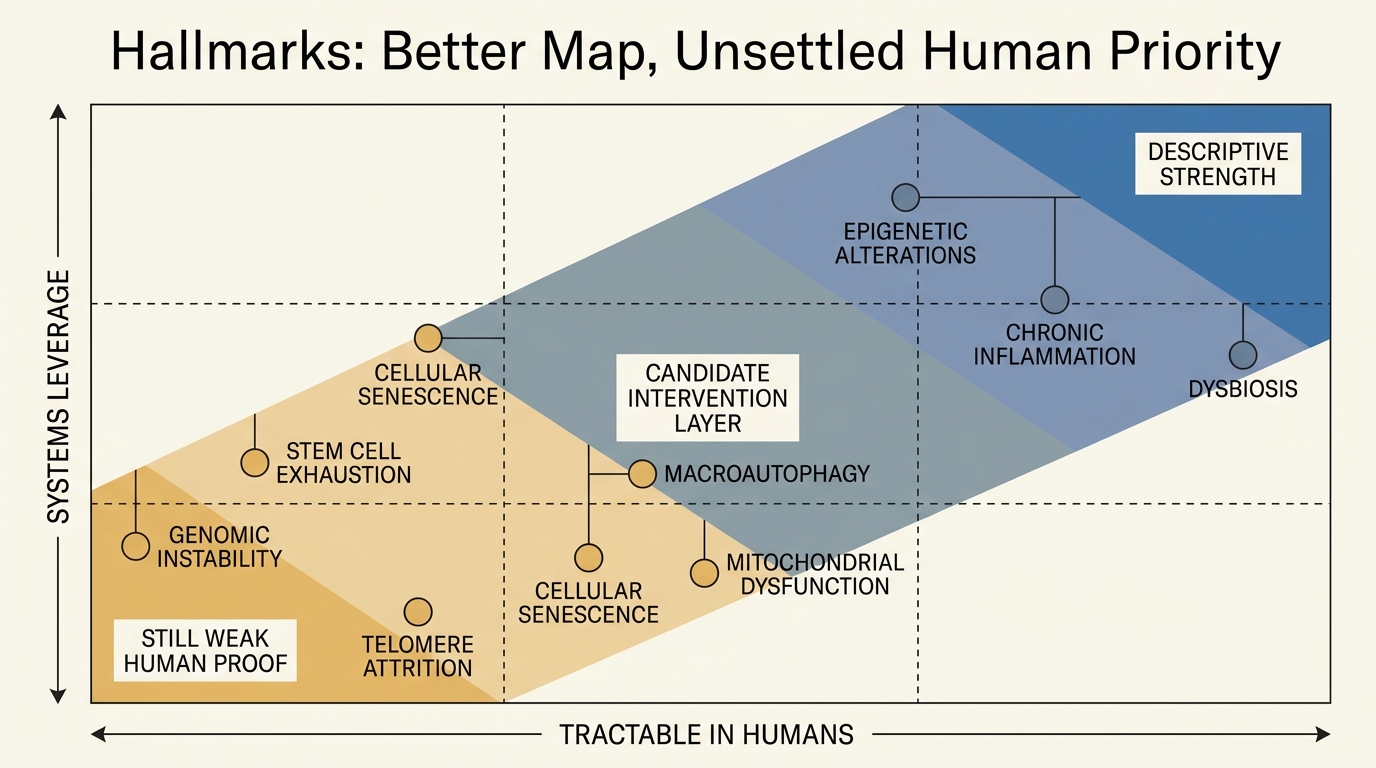
Hallmarks of Aging Revisited: What Changed Since 2013
A disciplined read of what the 2013 hallmarks paper established, what the 2023 expansion clarified, and why the framework still does not rank human leverage points cleanly.
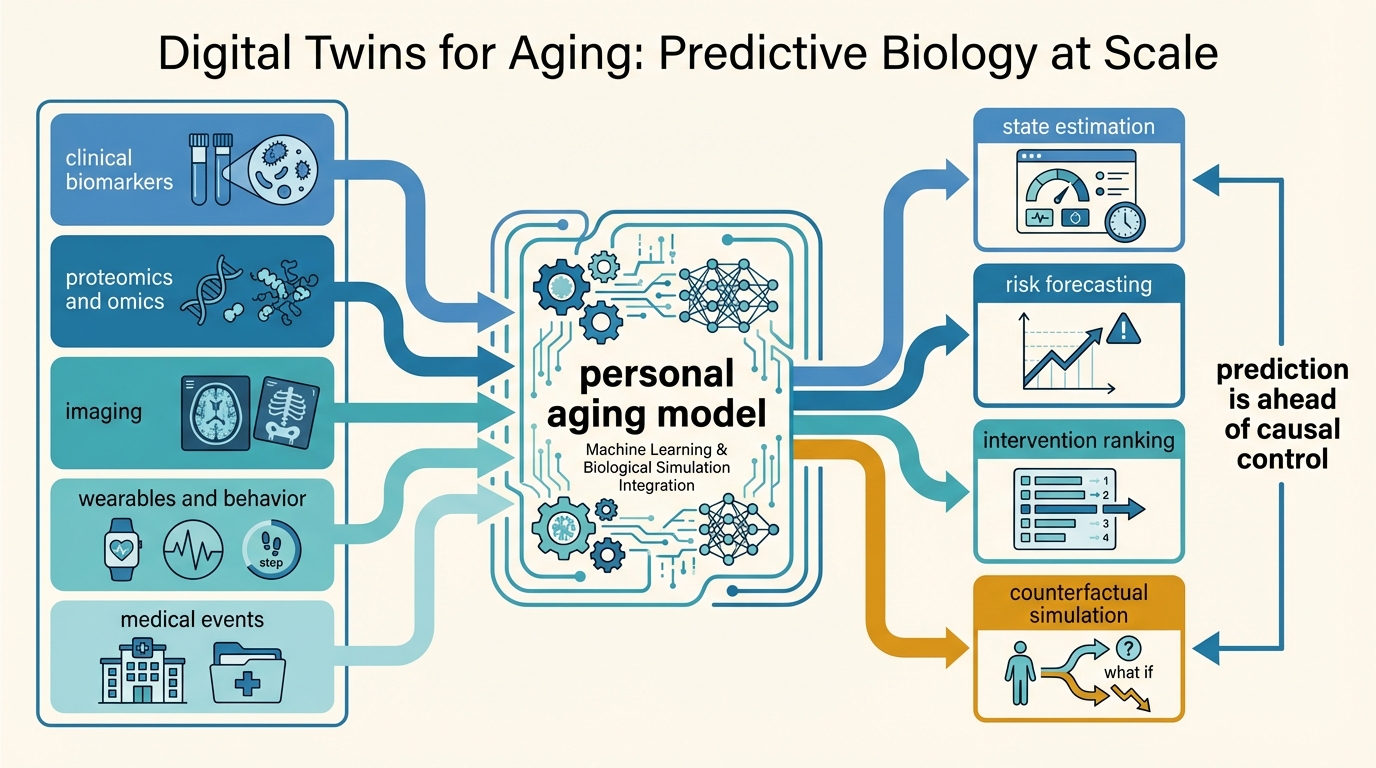
Digital Twins for Aging: Predictive Biology at Scale
A LifeMeter analysis of what aging digital twins can realistically do, why multimodal prediction is advancing faster than causal simulation, and where scale still outruns validation.
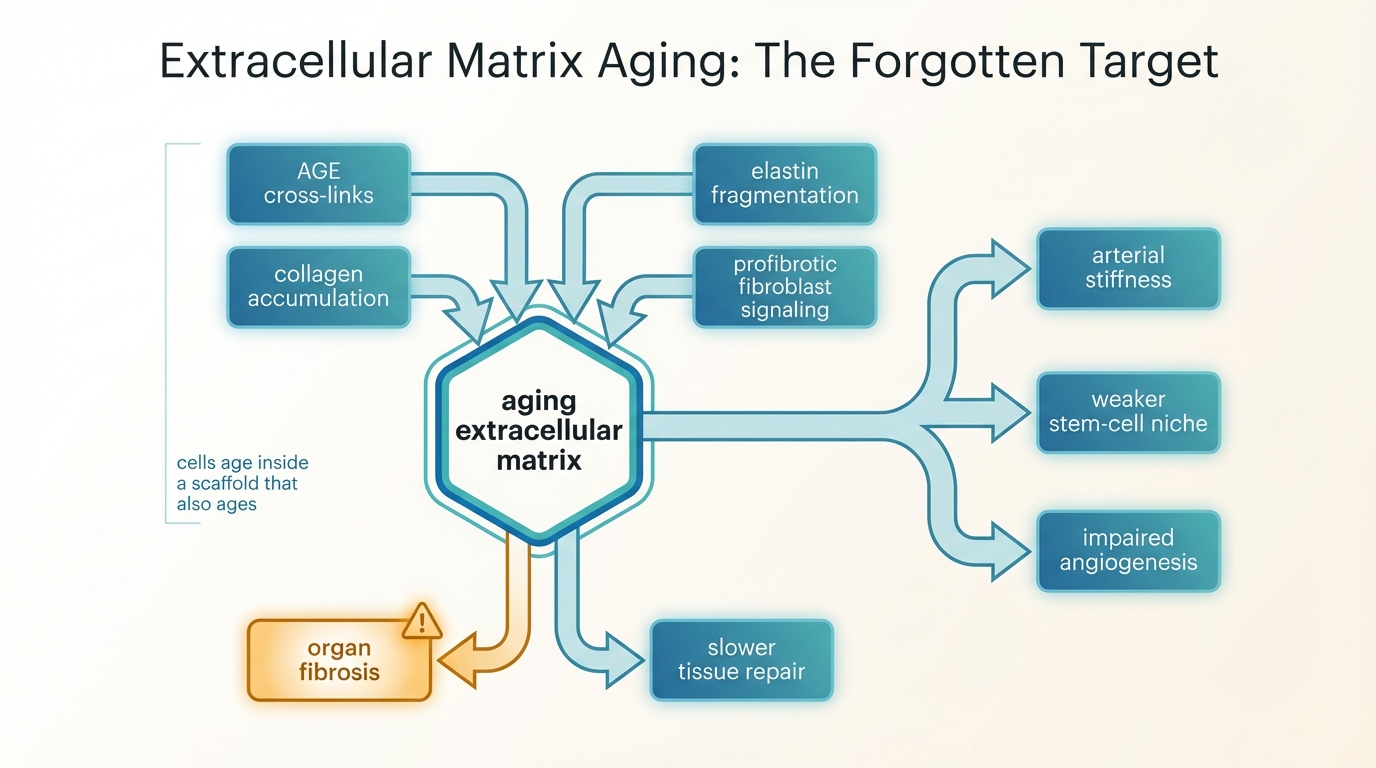
Extracellular Matrix Aging: The Forgotten Target
A LifeMeter analysis of why extracellular matrix aging matters, how collagen cross-linking and stiffness reshape tissue function, and why longevity medicine still underweights matrix repair.

Senomorphics vs Senolytics: Slowing vs Removing Damage
A LifeMeter audit of the difference between senomorphics and senolytics, where each strategy fits biologically, and why neither has earned blanket anti-aging use in humans.
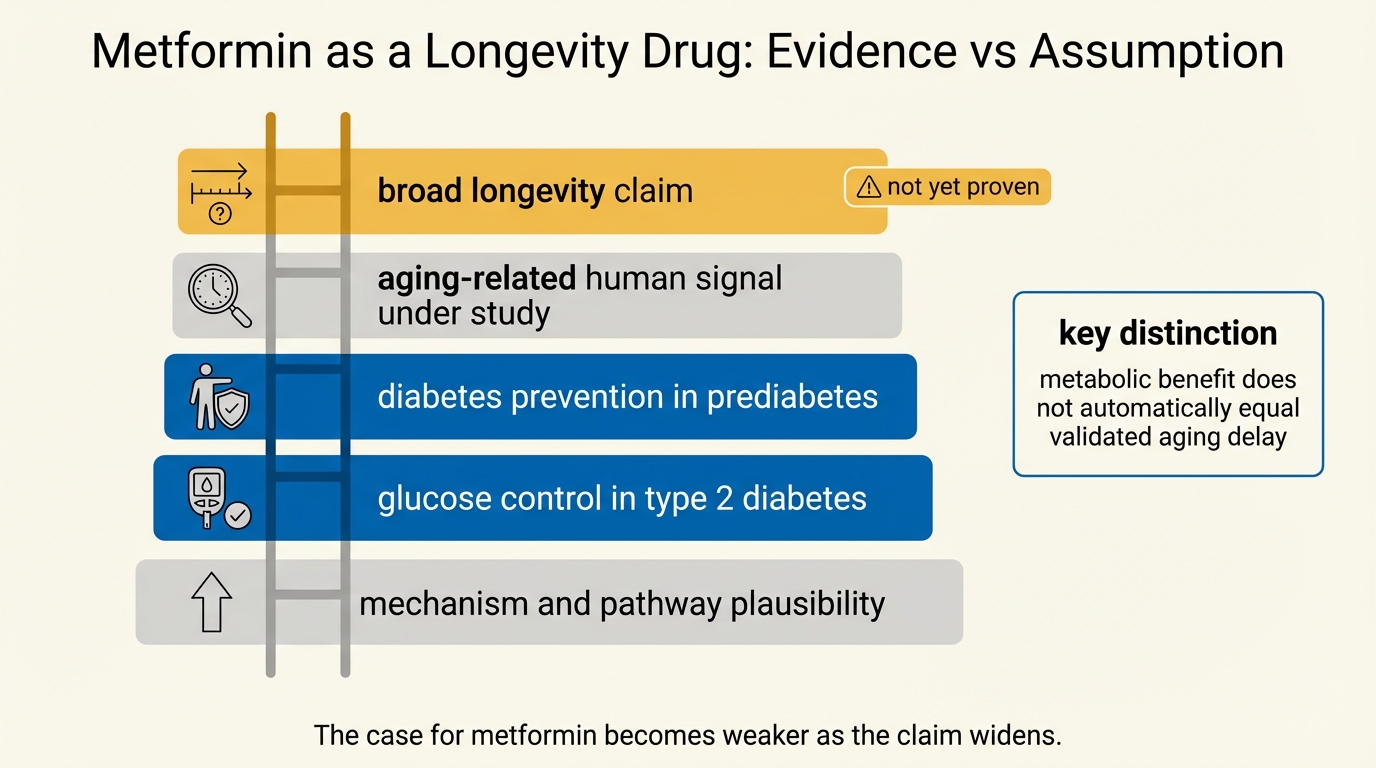
Metformin as a Longevity Drug: Evidence vs Assumption
A LifeMeter audit of metformin as a longevity candidate, separating established diabetes-prevention and safety evidence from the much weaker case for broad anti-aging use in otherwise healthy adults.
Metformin as a Longevity Drug: Evidence vs Assumption
An April 2026 audit of metformin as a longevity drug, separating what is established in diabetes care and geroscience from what remains assumption in healthy aging.
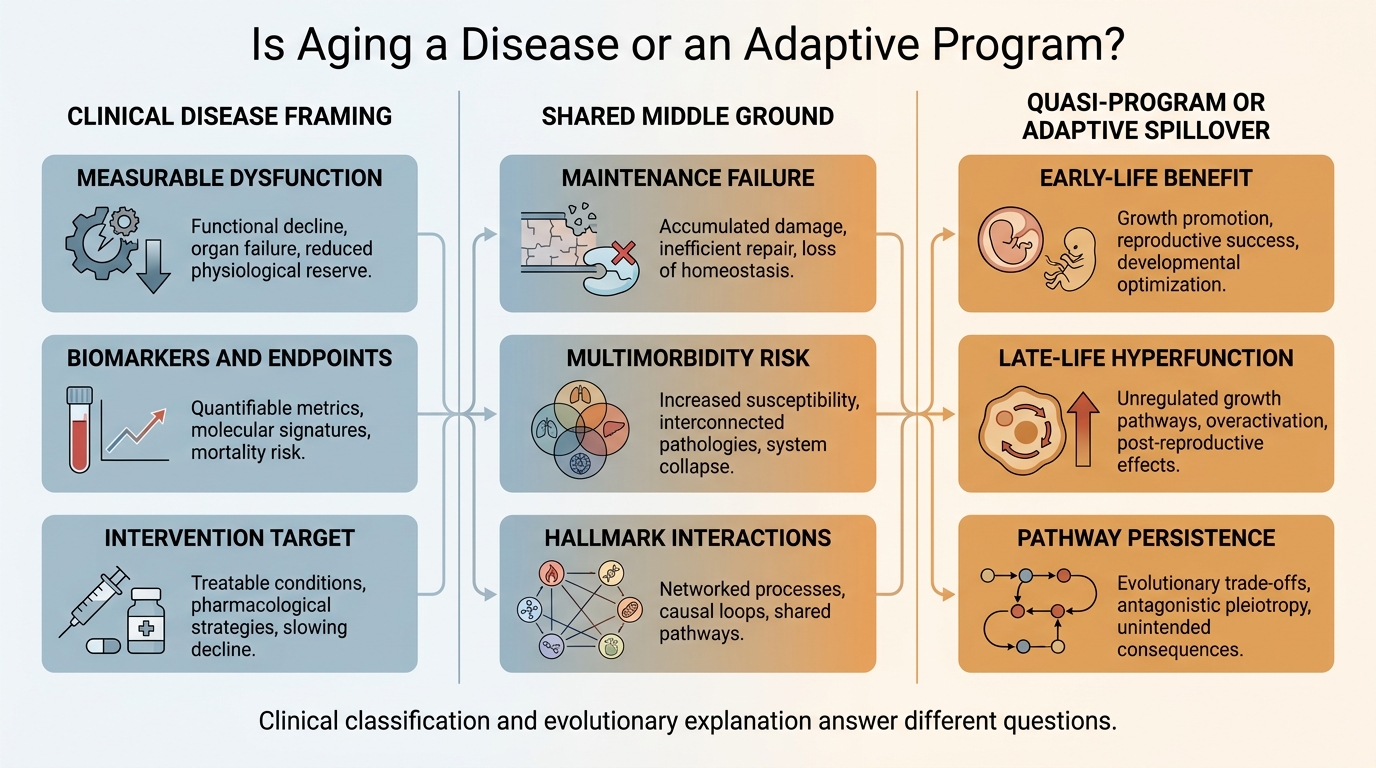
Is Aging a Disease or an Adaptive Program?
A 2026 refresh on the disease framing of aging, the quasi-program and adaptive-program debate, and why the strongest current view treats aging as a mixed process with different explanatory layers.
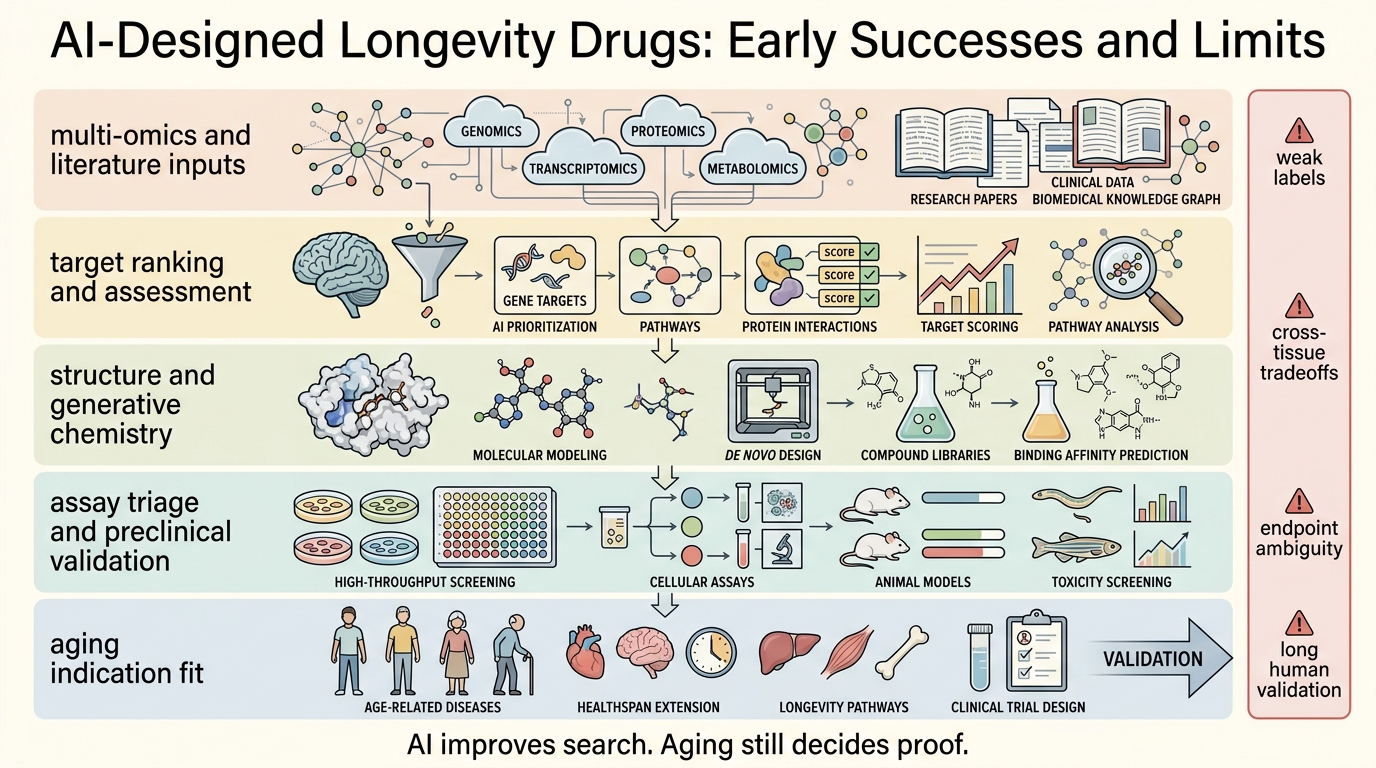
AI-Designed Longevity Drugs: Early Successes and Limits
A 2026 refresh on what AI has genuinely improved in longevity drug discovery, where aging programs still fail, and why faster models do not remove endpoint or validation risk.
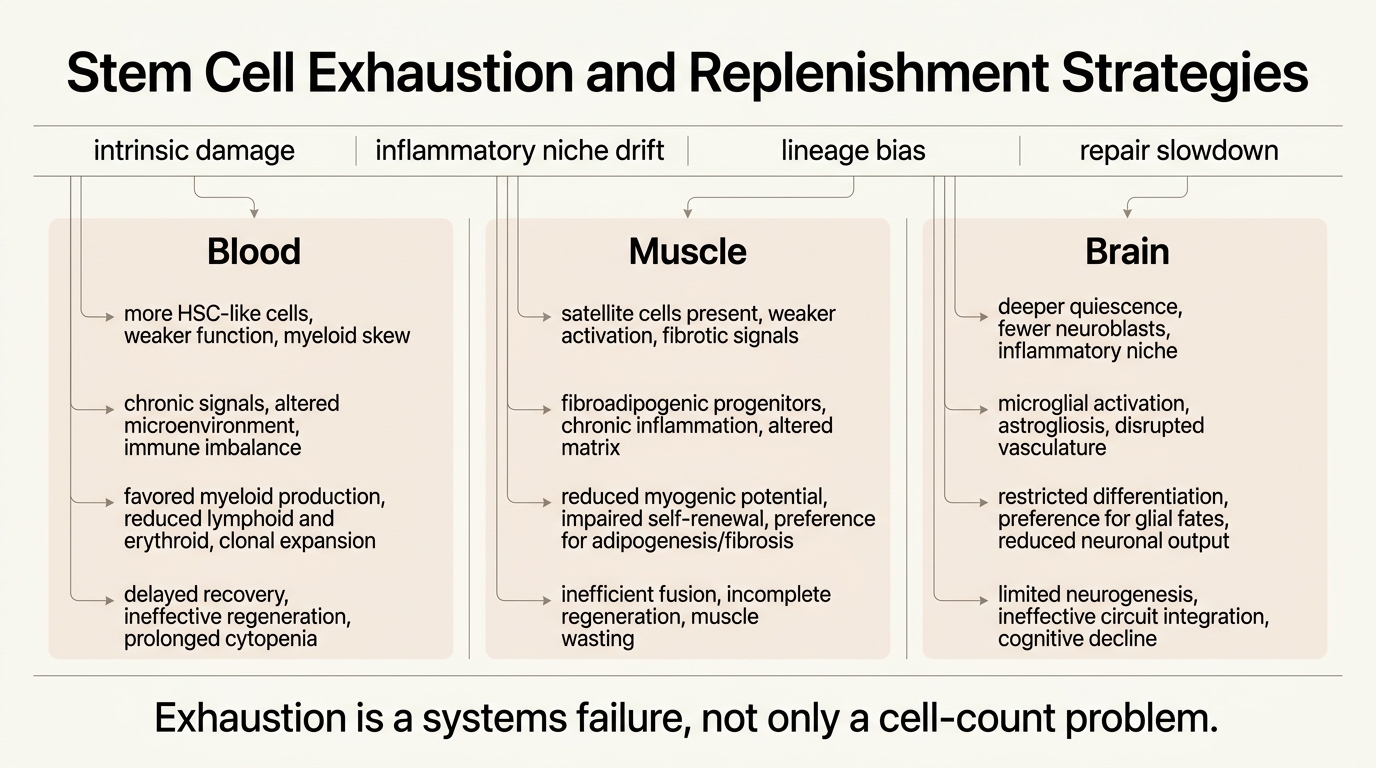
Stem Cell Exhaustion and Replenishment Strategies
A 2026 audit of stem cell exhaustion across blood, muscle, and brain, and why most realistic replenishment strategies aim to restore function and niche quality before they attempt wholesale replacement.

Caloric Restriction Mimetics: What Actually Works in Humans
An April 2026 audit of caloric restriction mimetics in humans, separating narrow clinical signal from the stronger claim of true calorie-restriction replacement.
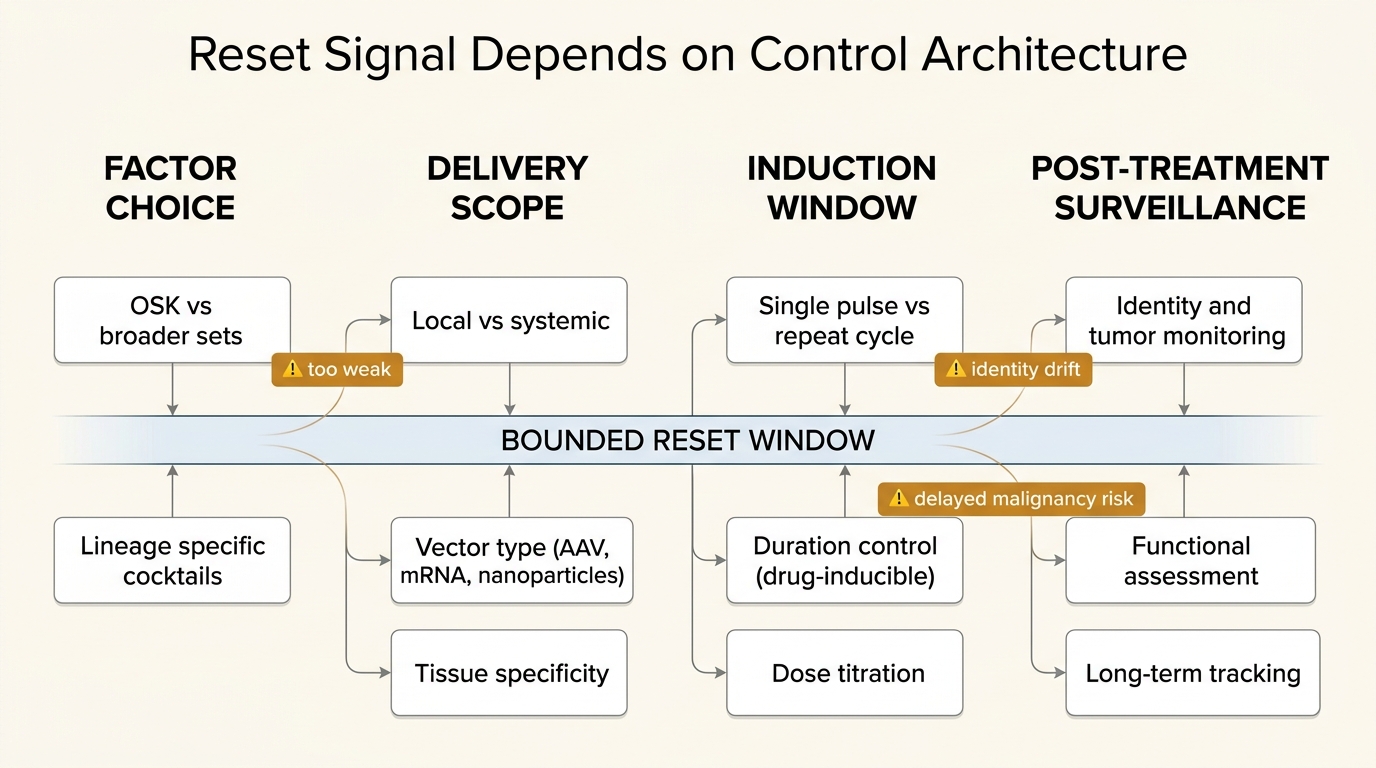
Partial Cellular Reprogramming: Reset Without Cancer Risk?
A disciplined evidence audit of partial cellular reprogramming, focused on what bounded in vivo studies established, where the 2026 skin data fits, and why cancer risk remains unresolved.
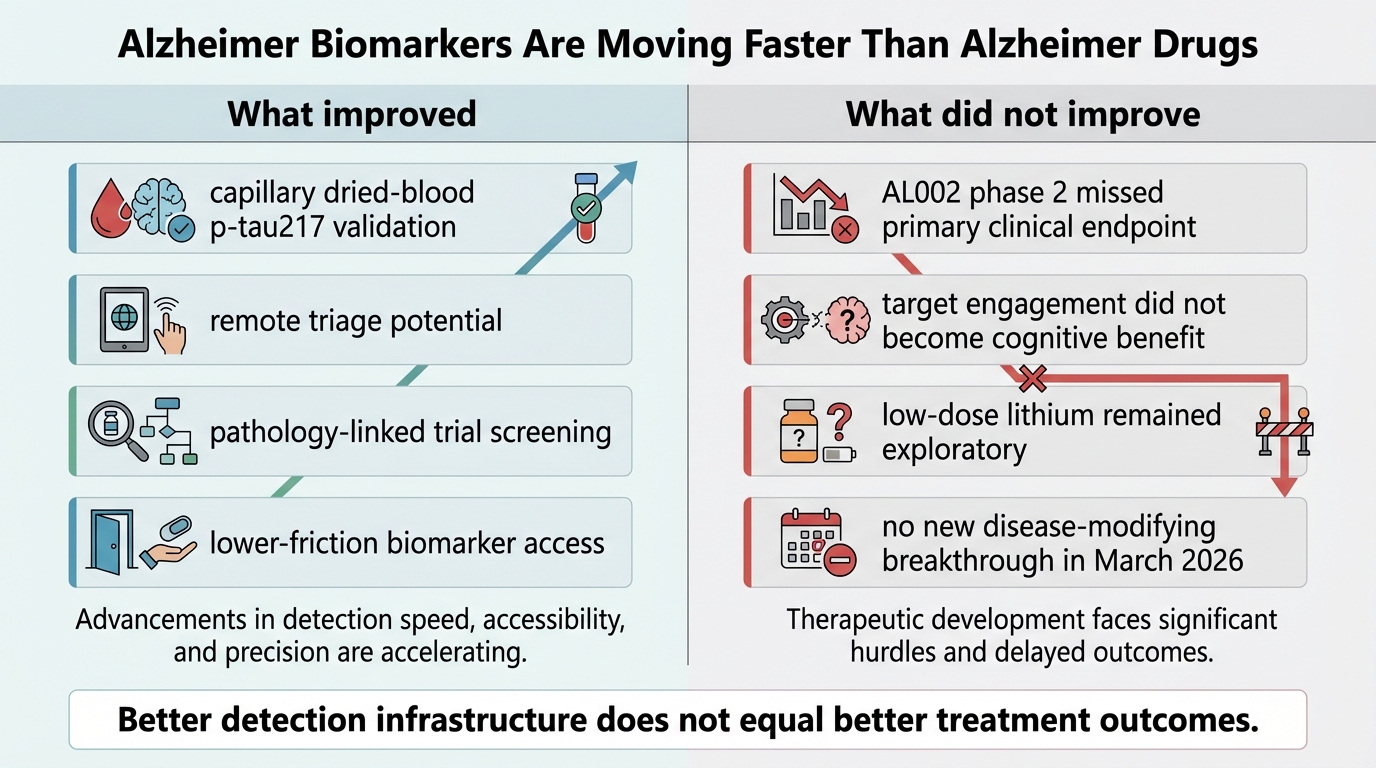
Alzheimer Biomarkers Are Moving Faster Than Alzheimer Drugs
March 2026 evidence review: Alzheimer blood biomarkers are scaling fast, but therapeutic progress remains slower and less certain.
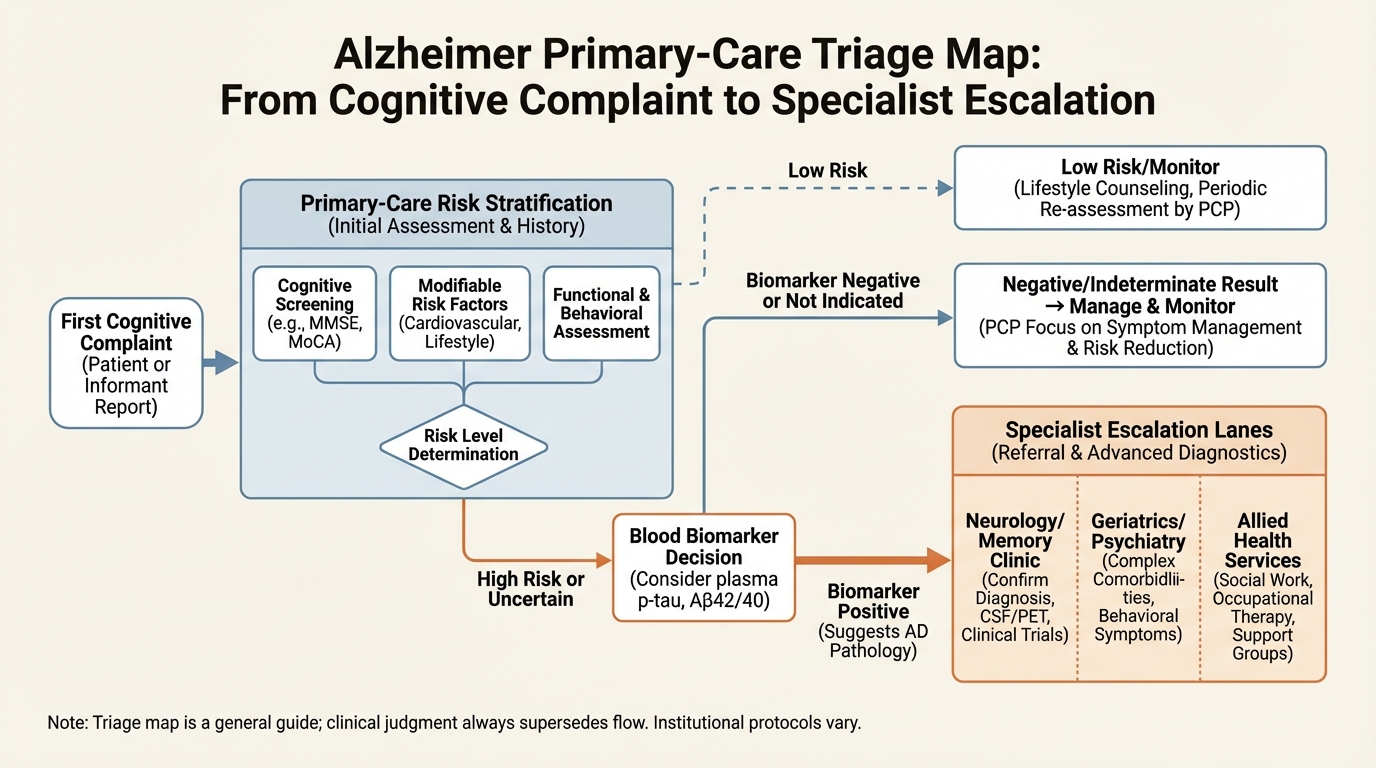
The Primary-Care Alzheimer's Pivot: What Blood Biomarkers Can and Cannot Do
How newly deployed blood biomarkers shift Alzheimer triage, referral timing, and family planning under uncertainty.
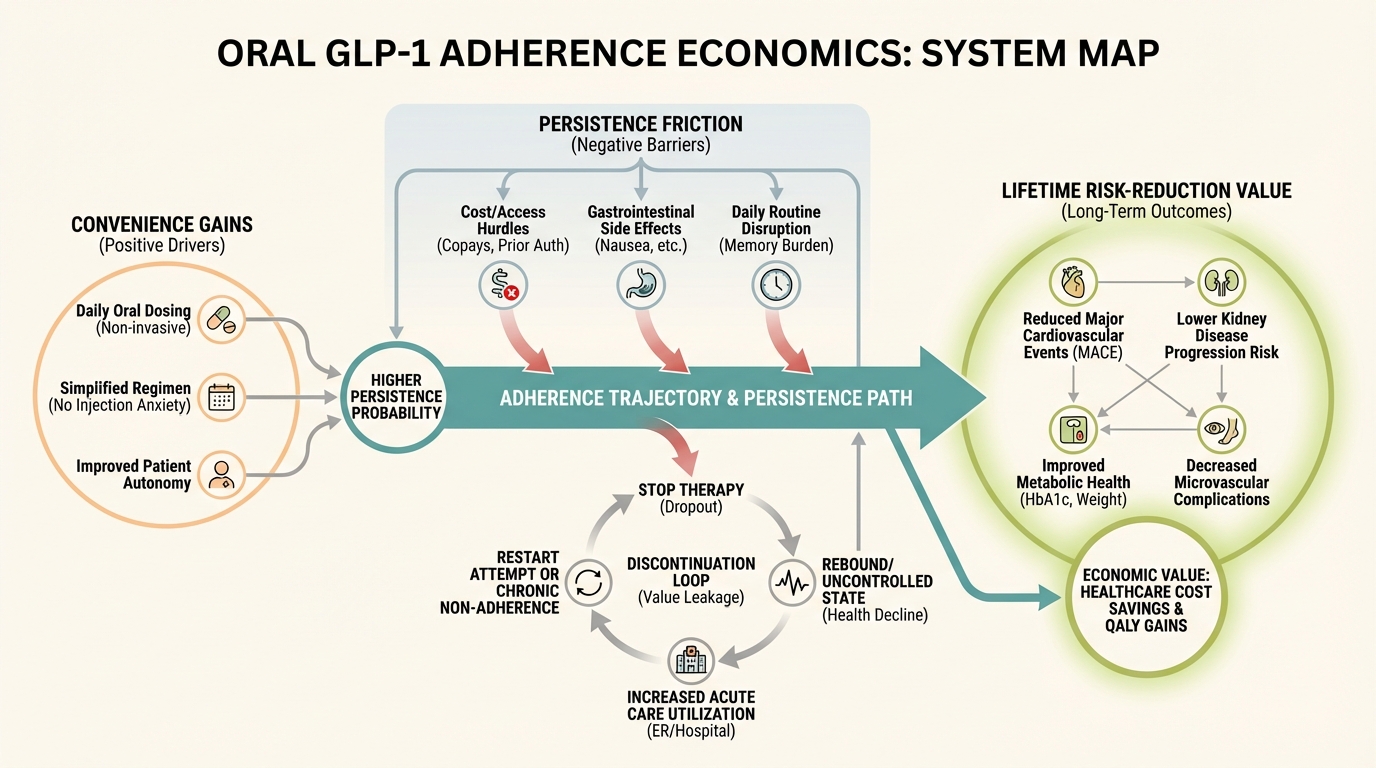
Oral GLP-1 Enters Phase 3 Reality: Convenience, Adherence, and Longevity Tradeoffs
What oral GLP-1 changes in adherence economics, discontinuation risk, and long-horizon cardiometabolic planning.
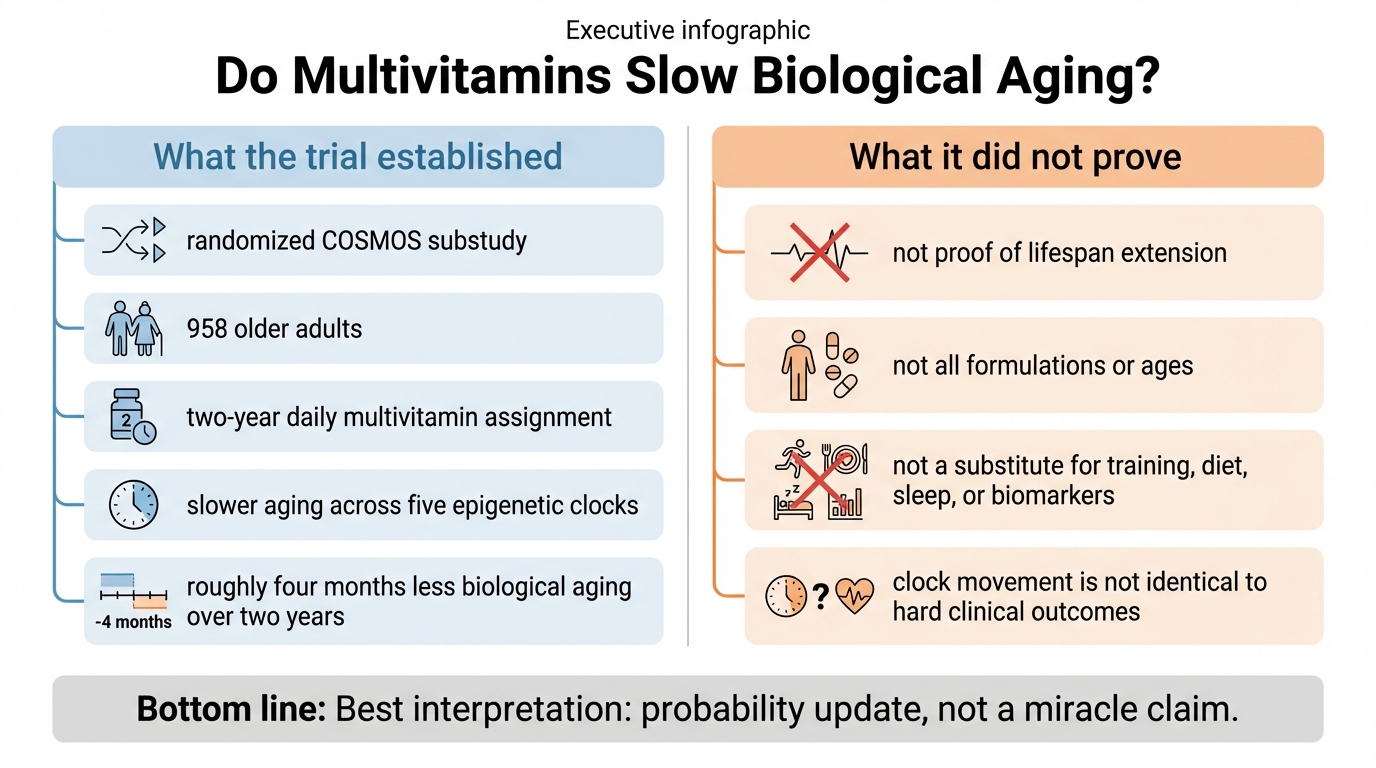
Do Multivitamins Slow Biological Aging? What the New COSMOS Clock Trial Shows, and What It Does Not
A precise read of the March 2026 COSMOS epigenetic-clock trial: what the multivitamin result establishes, what remains uncertain, and how to use it without self-deception.
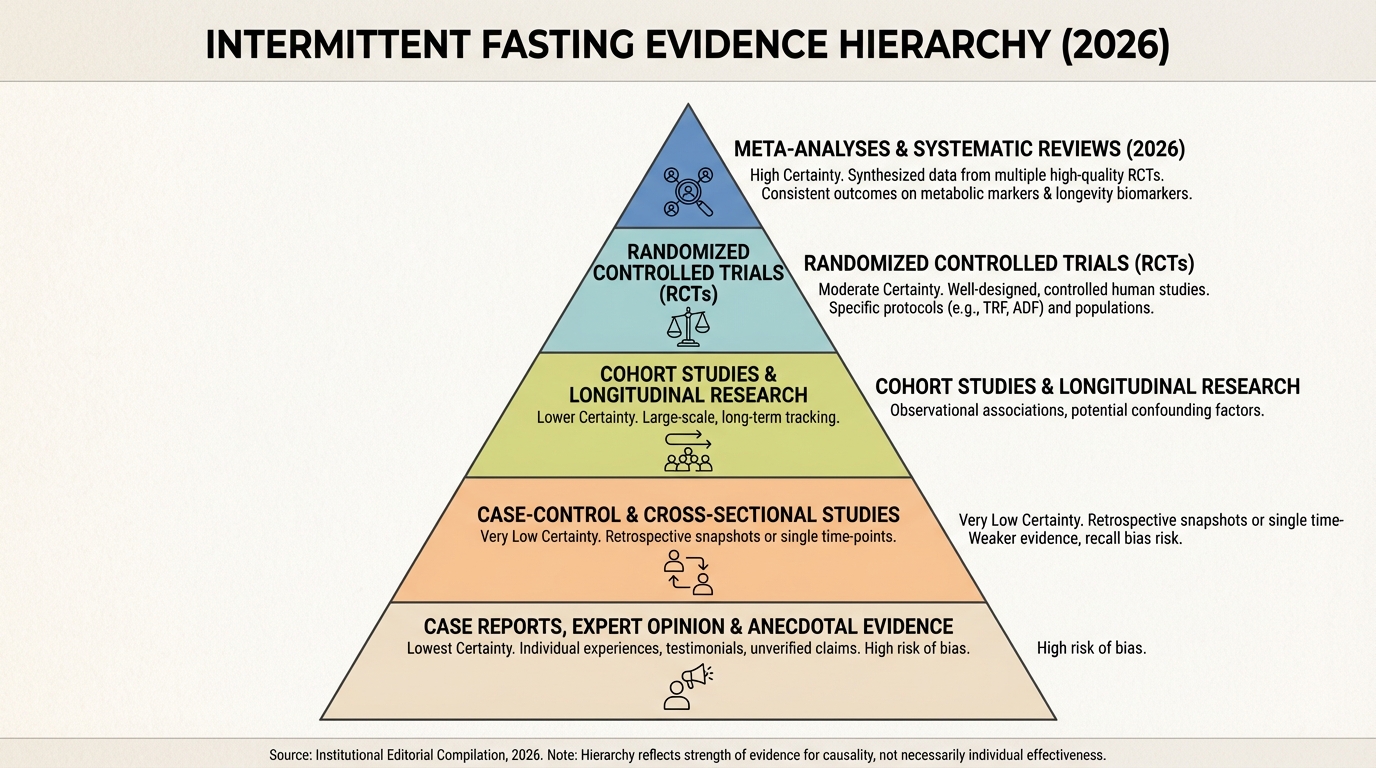
Intermittent Fasting After the Hype Cycle: What the 2026 Evidence Actually Supports
A practical reset after the 2026 evidence update: where fasting helps, where it does not, and how to decide quickly.
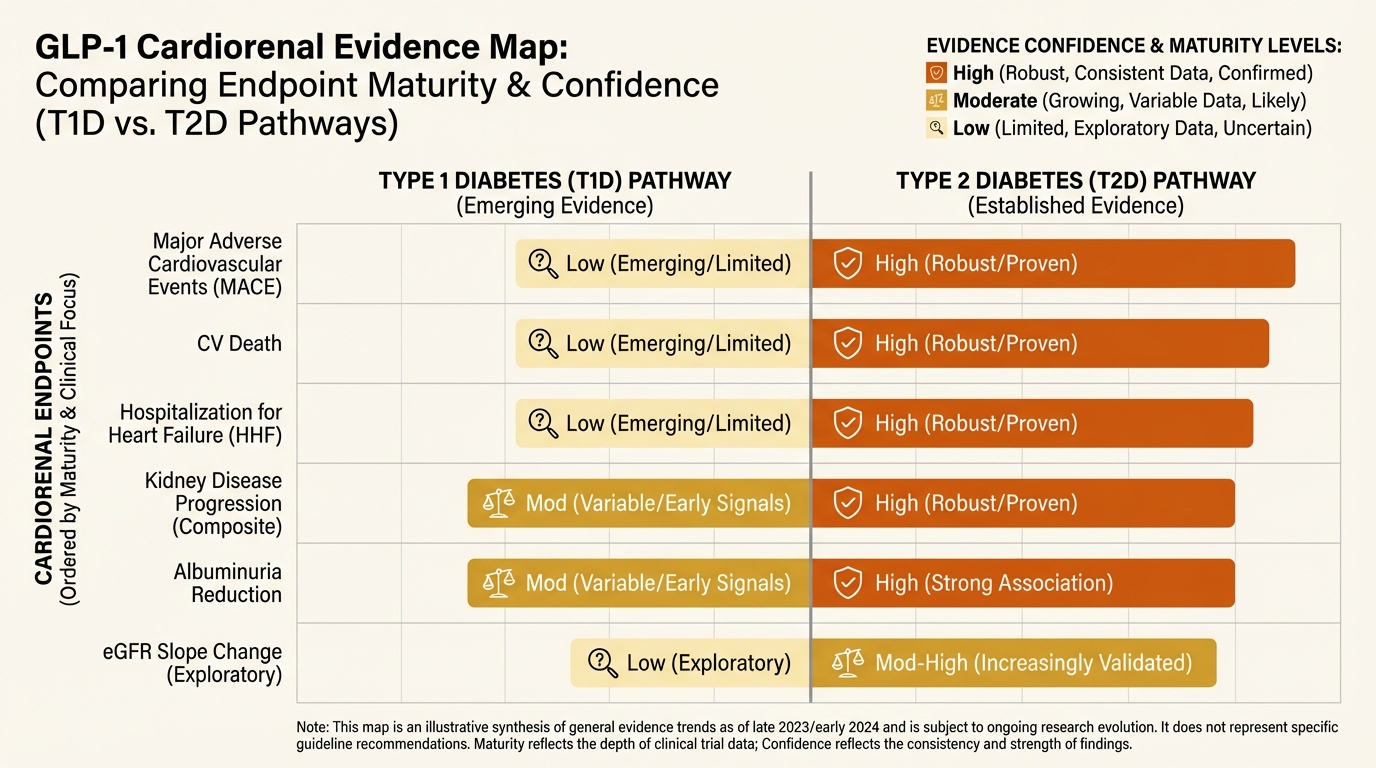
GLP-1 Beyond Type 2 Diabetes: Cardiorenal Signals, Safety Boundaries, and Off-Label Drift
How to read new cardiorenal GLP-1 signals without overextending into unsafe off-label practice.
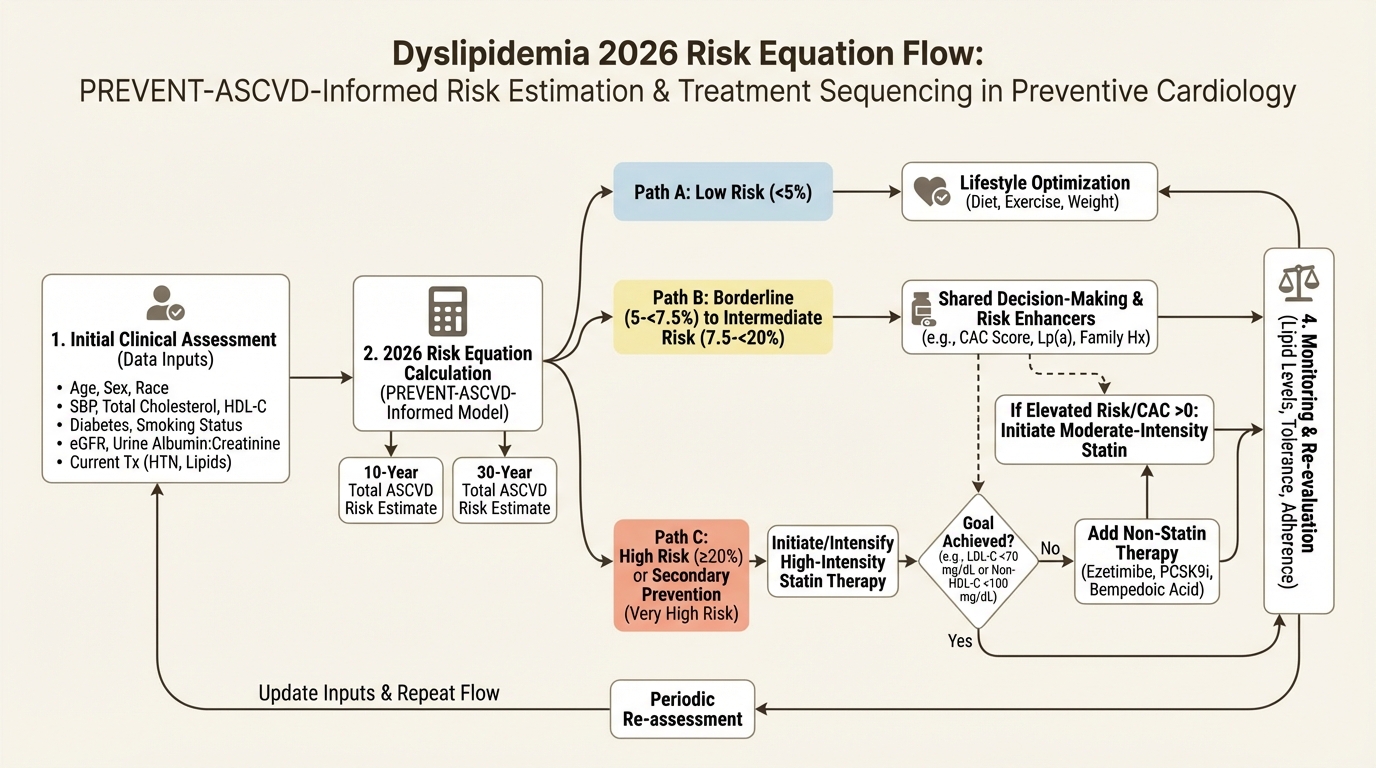
The 2026 Dyslipidemia Rewrite: Lp(a), ApoB, and 30-Year Risk in Preventive Longevity
How the 2026 dyslipidemia update changes prevention logic: PREVENT equations, apoB, Lp(a), CAC, and longer risk horizons.
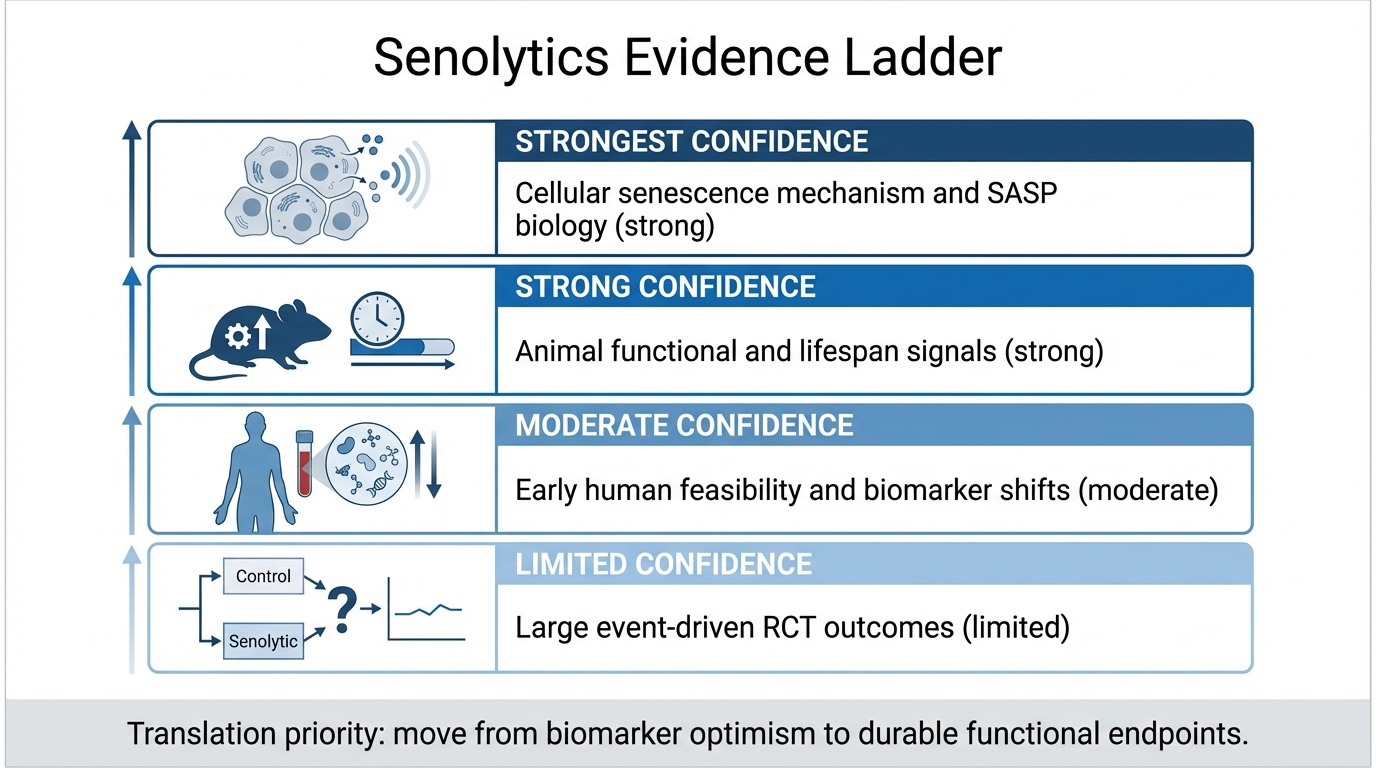
Senolytics: From Mouse Lifespan Gains to Human Trials
An April 2026 LifeMeter audit of senolytics, separating strong mouse evidence, early human target-engagement studies, and the missing clinical outcome proof.
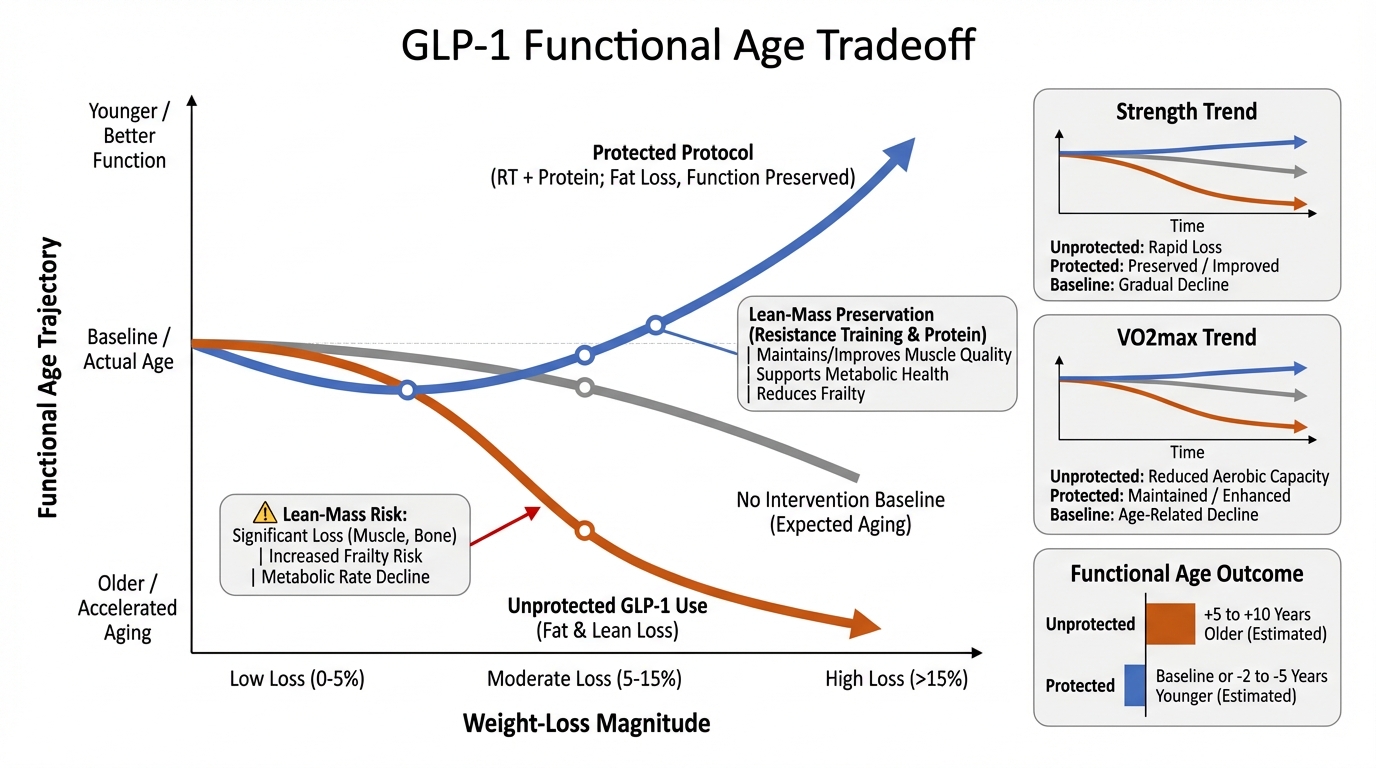
GLP-1 and Functional Age: When Weight Loss Helps Longevity and When It Hurts
A practical evidence review of GLP-1 and dual incretin therapy: metabolic gains, lean-mass risk, and the intervention stack required to protect functional age.
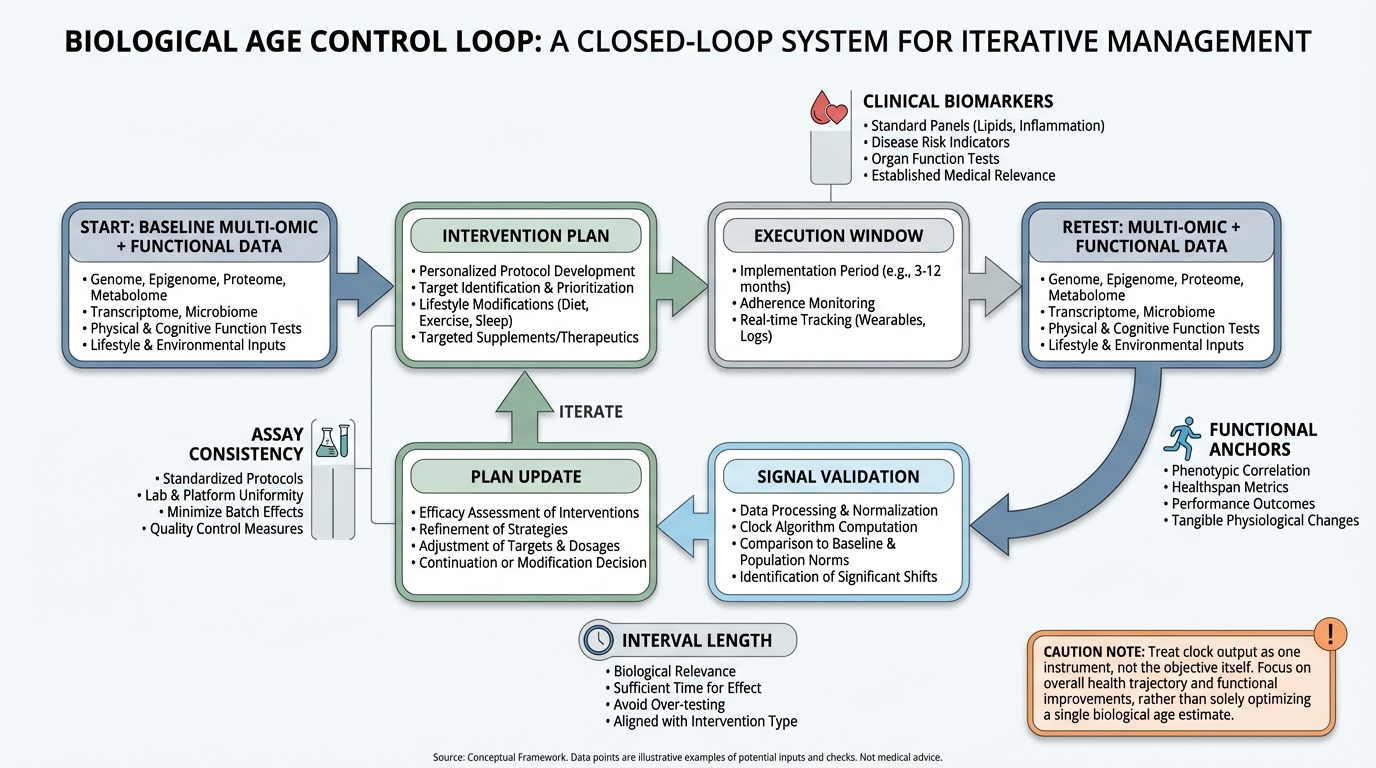
Biological Age Clocks as Decision Tools: What Is Real, What Is Overstated, and How to Use Them Without Self-Deception
A practical, evidence-graded guide to biological age clocks: what is validated, what remains noisy, and how to use clocks inside a real healthspan decision loop.
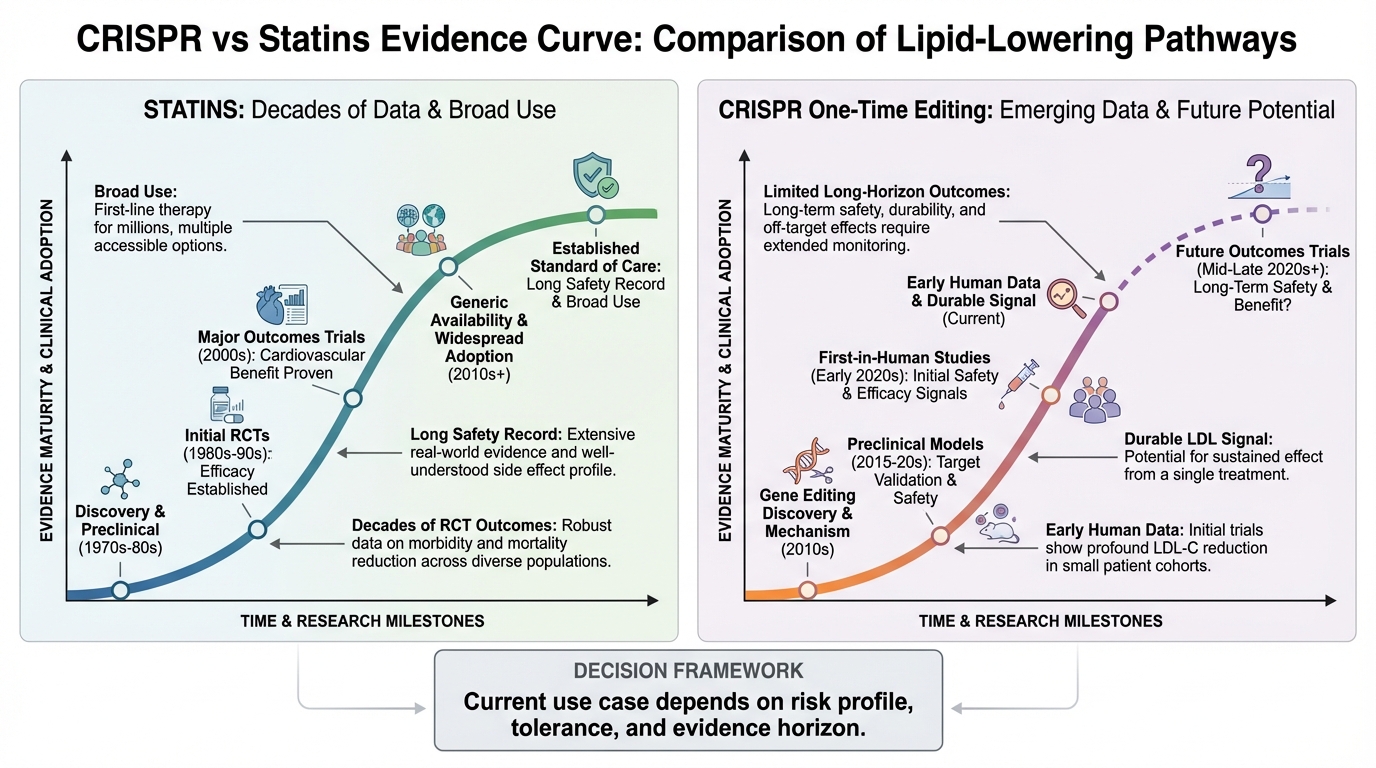
One Injection vs Lifetime Statins: What the CRISPR Cholesterol Trial Actually Shows
A practical breakdown of what the CTX310 Phase 1 CRISPR cholesterol data actually shows, where it is promising, and what remains unproven versus statin-standard care.
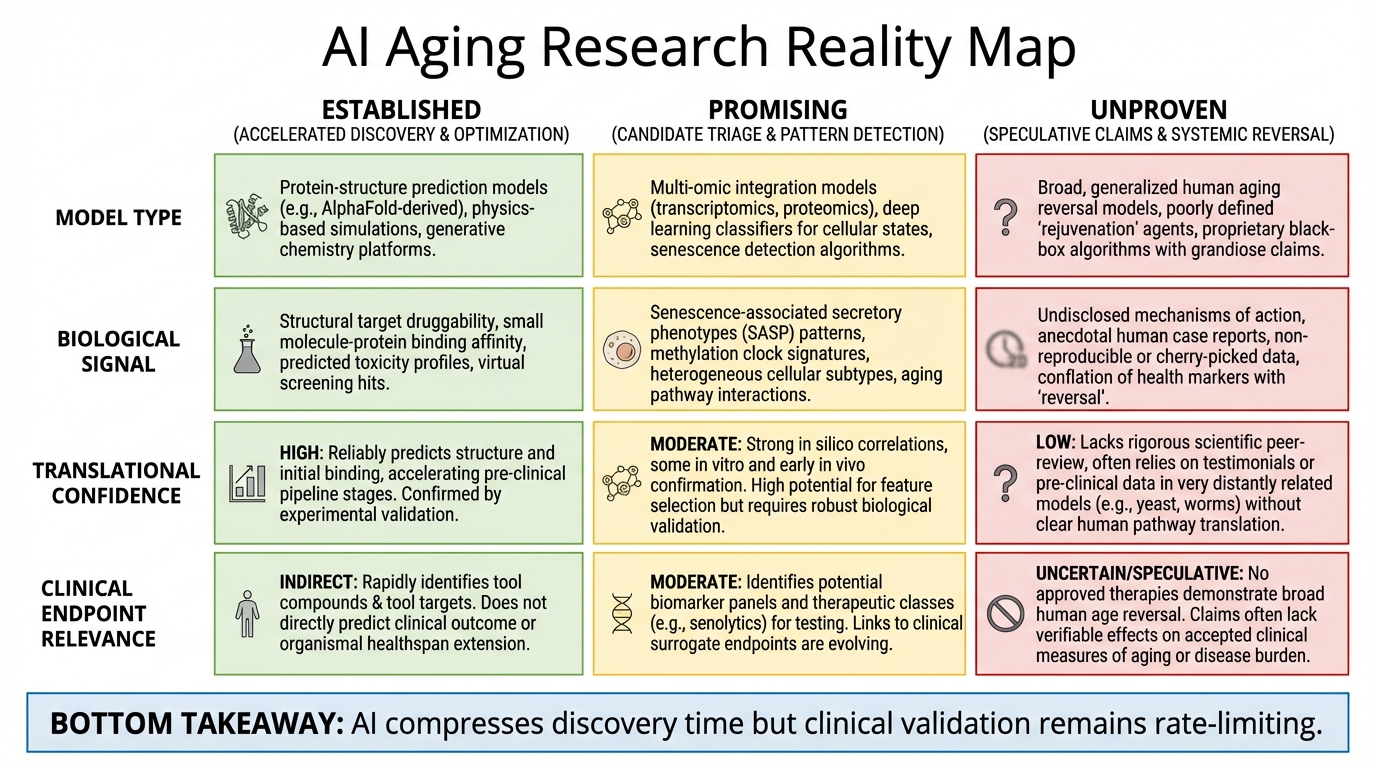
Did AI Reverse Aging? What Is Real, What Is Overstated, and What Comes Next
A clear evidence-based breakdown of AI and aging claims: what has been demonstrated in animals, what remains unproven in humans, and what likely comes next.

Tutorial 5: Biomarkers That Matter - Optimal Lab Values for Longevity and Healthspan
Master longevity biomarkers: optimal lab ranges for fasting glucose, HbA1c, ApoB, hs-CRP, and 20+ critical markers. Learn Tier 1, 2, and 3 testing protocols for healthspan extension.
Tutorial
Master financial planning for extended longevity. Learn the endowment model, multi-stage career framework, and capital strategies for 100+ year healthspan to avoid insolvency.
Tutorial
Understand the Death Gap: the distance between your biological state and the threshold to benefit from longevity therapies. Learn why functional capacity, not calendar age, determines survival.
Tutorial 2: Longevity Escape Velocity (LEV) Explained - When Medical Progress Outpaces Aging
Understand Longevity Escape Velocity (LEV): when medical science adds more than one year of life expectancy per year. Learn the timeline, mechanisms, and strategic implications for 2030-2045.
Tutorial
Learn the Bridge Strategy: a systematic framework to extend your healthspan through 2035 when breakthrough longevity therapies reach clinical availability. Master Tier 1, 2, and 3 interventions.