Tradeoffs in Longevity: Cancer Risk, Fertility, and Growth
People want the upside of youth without the liabilities that often come with strong growth biology. Longevity research keeps returning to the same constraint: the pathways that build tissue, support reproduction, and drive development early in life can also raise the long-run burden of cancer, hyperfunction, and repair imbalance later on.
The Boundary Most Readers Want to Ignore
Serious anti-aging work therefore sounds less like optimization and more like risk pricing. Lower growth signaling can improve lifespan and healthspan in many model systems. Yet humans are not mice raised for one endpoint. The same biology that can slow late-life deterioration also helps determine muscle and bone accretion, reproductive function, wound repair, and developmental timing.
Longevity strategy therefore has to separate a mechanistic insight from a usable human intervention. The insight is that persistent growth signaling can age tissues by favoring proliferation, suppressing cleanup programs, and widening cancer opportunity. The intervention question is narrower: how far can that signaling be reduced, at what age, in which tissue context, and at what cost to other functions that still matter?
Established fact: nutrient-sensing and growth pathways such as GH-IGF-1 and mTOR influence development, metabolism, cancer biology, and aging across many species. Reasoned inference: the strongest longevity gains are unlikely to come from maximizing or minimizing those pathways universally, but from timing and dose patterns that reduce late-life damage without collapsing necessary human function.
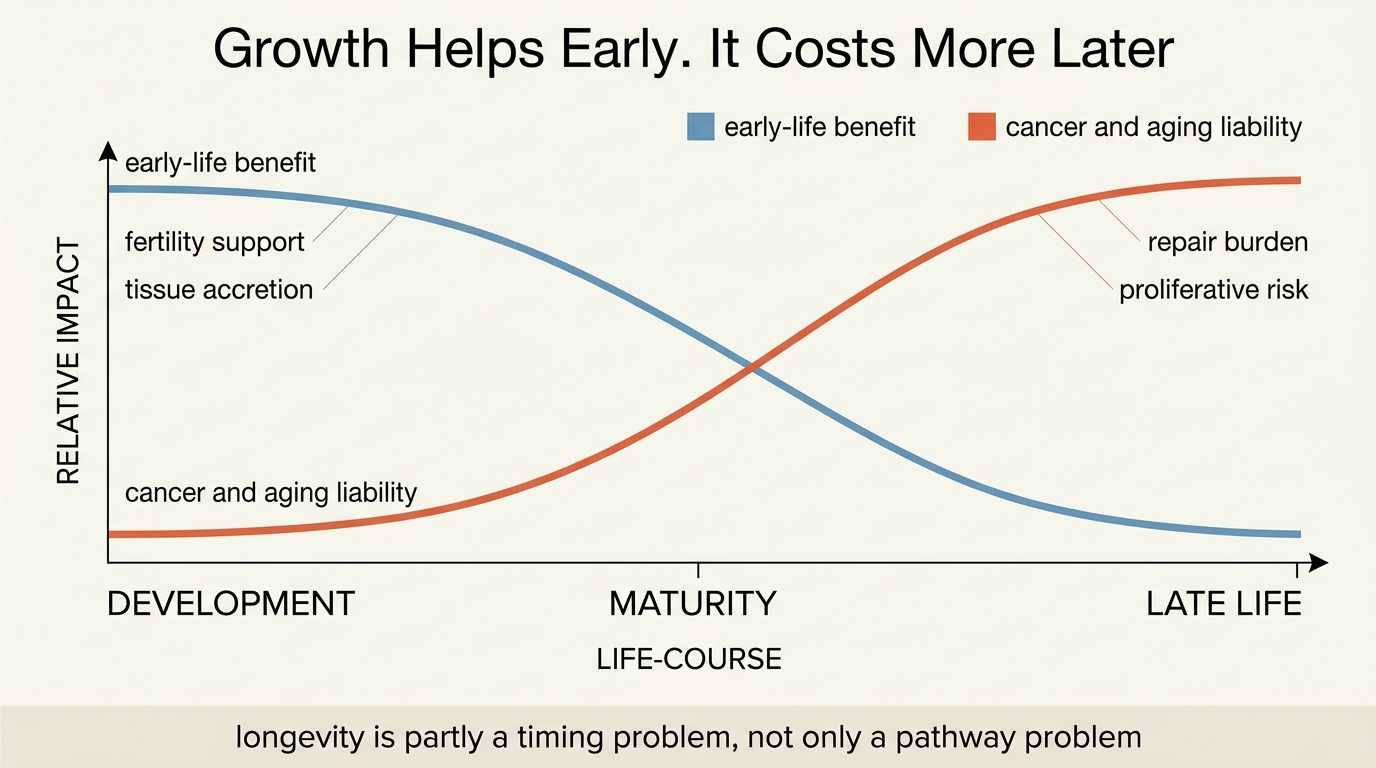
Why Growth Becomes a Longevity Problem
Growth is expensive. It asks cells to divide, synthesize protein, expand tissue, and keep nutrient throughput high. Those functions are useful when an organism is building itself, reproducing, or recovering from acute damage. They become less clean later in life because the cost of continued proliferation rises when DNA damage, senescent burden, clonal drift, and tissue heterogeneity have already accumulated.
This is why longevity biology repeatedly returns to mTOR, insulin signaling, and IGF-1. These pathways do not merely correlate with aging. They help govern how strongly the organism prioritizes building over maintenance. In many models, dialing that pressure down extends lifespan because autophagy, stress resistance, and proteostatic discipline improve when the system stops treating every phase of life like a growth phase.
Antagonistic Pleiotropy Is a Better Frame Than Miracle Pathways
The evolutionary language here is useful because it prevents false promises. Antagonistic pleiotropy means a trait can help early-life fitness while harming late-life survival. That logic fits growth programs unusually well. A system selected to support faster development, larger body size, or stronger reproductive output does not have to be optimal for surviving deep into old age. Natural selection discounts late-life costs once reproduction has largely been secured.
This does not reduce every longevity claim to one theory. It does mean there is no free lunch. If a pathway increases early-life reproductive success, tissue growth, or competitive advantage, there is a serious prior that it may also carry later-life tradeoffs in cancer exposure, metabolic strain, or repair quality.
The GH-IGF-1 and mTOR Tradeoff Stack
The GH-IGF-1 axis and mTOR are not identical, but they belong in the same practical frame because both sit close to the decision boundary between growth and maintenance. Higher activity supports anabolism and tissue-building. Lower or more restrained activity often improves stress resistance and can reduce proliferative pressure. These pathways therefore matter in both oncology and geroscience.
Translation is conditional. A frail older adult is not helped by a stripped-down longevity model that ignores sarcopenia, fracture risk, immune competence, or healing after illness. An intervention that looks anti-aging in one context can become deconditioning in another if it pushes too far against necessary anabolic support. The question is not whether less signaling can help. It is when the tradeoff turns from protective restraint into underpowered physiology.
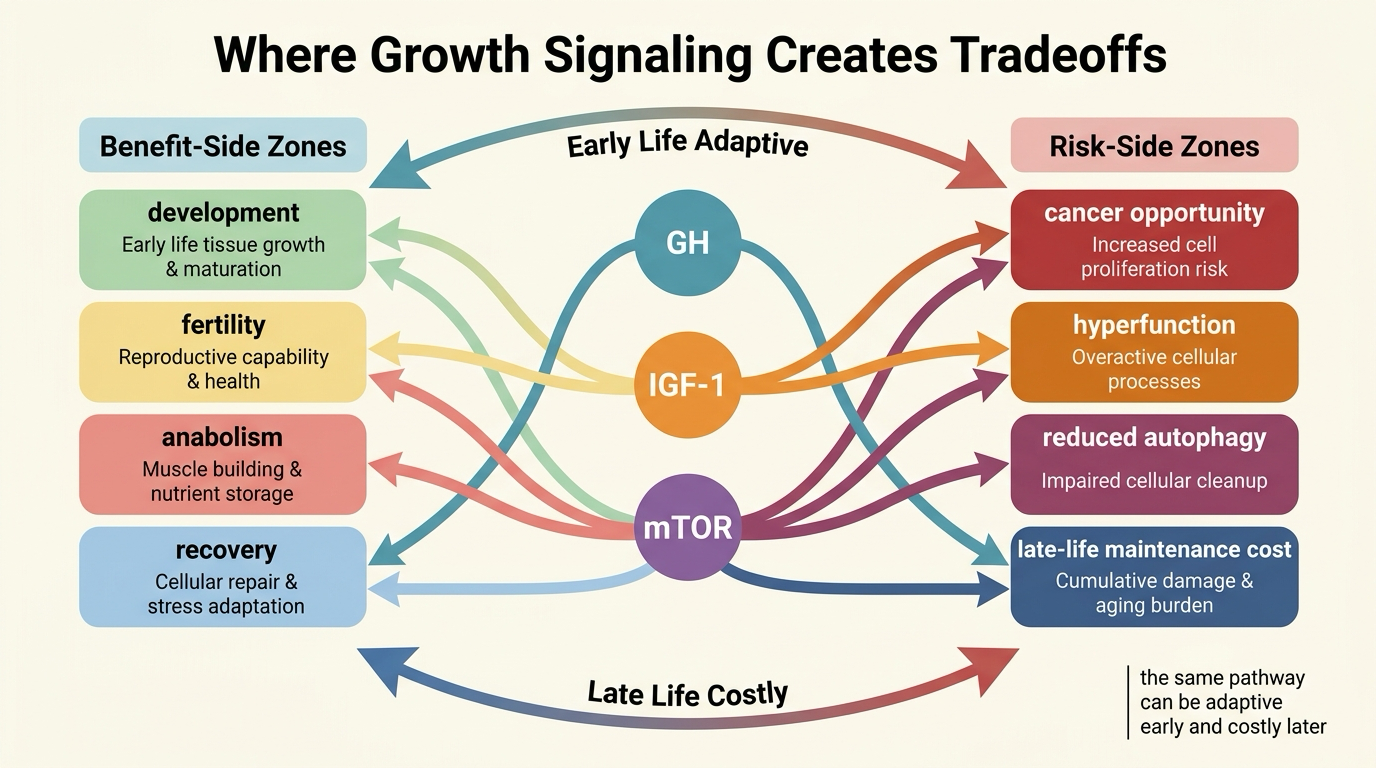
Fertility Is Not a Side Note
Fertility is where many simplified longevity narratives break down. Reproductive biology is tightly linked to energy availability, endocrine signaling, and organismal readiness to invest in offspring. Interventions that suppress growth signaling, lower nutrient throughput, or push chronic scarcity cues can reduce fertility even when they improve lifespan in model organisms.
This matters because it exposes the limit of using lifespan extension as the only objective. A molecule or diet pattern that improves one endpoint in a controlled model can still be unattractive for humans if the cost is impaired reproductive function, menstrual disruption, poor recovery, or chronic underfueling. That does not weaken the biology. It clarifies the human use case.
| Question | Higher growth emphasis | Higher maintenance emphasis |
|---|---|---|
| Early-life outcome | Supports development, reproduction, and tissue accretion | Often slows growth or reduces reproductive drive under constraint |
| Late-life risk | Can widen cancer and hyperfunction pressure | Can improve stress resistance and cleanup capacity |
| Human danger | Overgrowth, metabolic strain, proliferative burden | Frailty, under-recovery, endocrine disruption if pushed too far |
| Decision frame | Do not treat more as automatically healthier | Do not treat less as automatically better either |
Human Translation Is About Timing, Not Purity
The practical implication is that age, baseline health, and objective matter more than doctrine. A younger adult trying to preserve fertility, strength, and performance should not read animal lifespan data the same way as an older adult trying to reduce cancer risk and metabolic overdrive after peak reproductive years. The optimal point on the curve is unlikely to be fixed across life stages.
That is also why interventions such as amino acid restriction, insulin-sensitivity work, and cautious pathway-targeting drugs have to be judged by functional outcomes, not by ideological commitment to “less growth.” Long-lived biology is not the same as chronically suppressed biology.
Decision-Grade Take for Readers
The right takeaway is not that longevity requires permanent deprivation. It is that organisms pay for growth. Early-life building, reproductive capacity, and tissue turnover do not come free. Anti-aging medicine becomes more credible when it admits that tradeoff openly and asks where the burden can be shifted without breaking the functions people still need.
- Treat growth pathways as context-dependent controls, not as villains or saviors.
- Expect cancer, fertility, muscle, and repair to remain linked in any honest longevity strategy.
- Prefer interventions that show functional benefit without demanding chronic undernourishment or endocrine destabilization.
- Be skeptical of any anti-aging claim that promises lifespan upside with no tradeoff discussion at all.
Known, Inferred, and Unknown
| Category | Assessment |
|---|---|
| Known | mTOR and GH-IGF-1 signaling influence growth, metabolism, reproduction, cancer biology, and aging across species. |
| Known | Reduced growth signaling often extends lifespan in model organisms, but the accompanying physiological costs vary by species, sex, tissue, and intervention method. |
| Known | Fertility and reproductive biology are sensitive to nutrient and growth signaling, which means lifespan-oriented interventions can impose reproductive costs. |
| Inferred | The most usable human longevity strategies will likely rely on phase-specific restraint rather than permanent maximal suppression of anabolic signaling. |
| Unknown | How far humans can safely modulate growth pathways over decades before the balance shifts from anti-aging benefit toward frailty, poor recovery, or endocrine harm. |
Source Frame
This article uses the broad pathway review mTOR at the nexus of nutrition, growth, ageing and disease for the link between nutrient sensing, cellular growth, and late-life maintenance costs.
For the aging-cancer axis, it draws on The key role of growth hormone-insulin-IGF-1 signaling in aging and cancer, which frames why growth signaling can be protective in one domain and destabilizing in another.
For the evolutionary tradeoff lens, it uses Is antagonistic pleiotropy ubiquitous in aging biology?. For fertility-specific pathway costs, it uses Emerging Role for Mammalian Target of Rapamycin in Male Fertility. The open question is not whether tradeoffs exist. It is how precisely human medicine can move them.
Translate this longevity claim into a capital-runway decision.
Life extension logic only matters if the balance sheet can carry it. Move into WealthMeter to compare assets, spending, and yield assumptions against the same long-horizon planning problem.