Extracellular Matrix Aging: The Forgotten Target
Longevity medicine usually tells an intracellular story. It tracks damaged DNA, mitochondrial decline, senescent cells, and stem-cell exhaustion. Those are real targets. Tissue function still depends on the scaffold surrounding those cells. When the extracellular matrix stiffens, cross-links, fragments, calcifies, or shifts its signaling state, repair quality degrades even if the cells themselves improve.
Established fact: the extracellular matrix, or ECM, is not inert packing material. It is a mechanically active and biochemically active tissue environment composed of collagens, elastin, proteoglycans, glycoproteins, basement-membrane components, and matrix-bound signaling factors. Established fact: aging changes matrix composition, cross-link density, stiffness, and turnover across multiple organs including vasculature, heart, lung, skin, skeletal muscle, and kidney. Established fact: those changes alter stem-cell behavior, angiogenesis, immune trafficking, mechanotransduction, and organ compliance. The practical implication is straightforward. A therapy that improves cells while leaving an aged matrix untouched may hit a structural ceiling.
Core thesis: extracellular matrix aging is a central and under-targeted driver of tissue decline because it changes both mechanics and signaling. Cell-focused rejuvenation strategies will often underperform if the surrounding scaffold remains stiff, fibrotic, cross-linked, and biologically misinformative. The field has stronger evidence for the problem than for any general solution, which is why matrix repair belongs in serious longevity analysis but not yet in blanket intervention claims.
Why The Matrix Matters More Than Most Longevity Narratives Admit
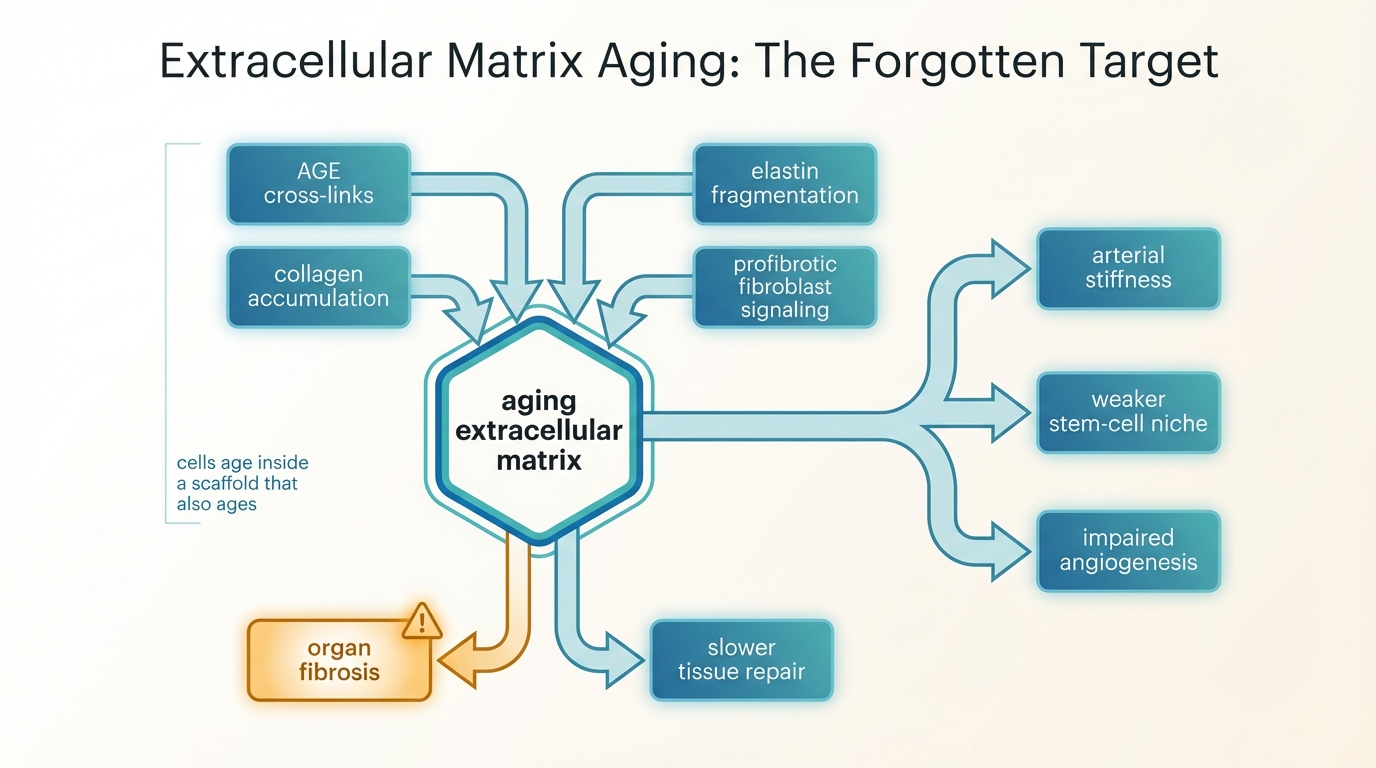
The ECM determines far more than structural support. It sets local stiffness, anchors cells through integrins and other receptors, stores growth factors, shapes diffusion, and constrains how tissues bear load. In a young tissue this scaffold is dynamic but regulated. In an old tissue the same scaffold can become an active source of distortion. Collagen accumulates and becomes more cross-linked. Elastin fragments and loses recoil. Fibrotic remodeling replaces compliant architecture with denser and less reversible material. The result is not one defect. It is a change in the rules under which cells operate.
That matters because cells read their environment constantly. Mesenchymal stem cells, endothelial cells, fibroblasts, immune cells, and muscle satellite cells respond to stiffness and matrix composition through mechanosensitive pathways such as integrin signaling, YAP and TAZ activity, focal adhesion dynamics, and cytoskeletal tension. Aged matrix can therefore push cells toward less regenerative and more inflammatory behavior even when those cells remain partly competent. This is one reason a tissue can stay old after a seemingly promising cell intervention.
It also explains a recurring translational failure mode. Preclinical work often treats tissue decline as though the main problem sits inside the target cell population. In reality the aged tissue niche includes matrix damage, altered fibroblast behavior, impaired vascular support, and inflammatory feedback loops. If those elements remain in place, cell replacement or reprogramming may produce weaker and less durable effects than the headline mechanism suggests.
What Matrix Aging Looks Like Biologically
The first axis is cross-linking. Long-lived proteins such as collagen and elastin are exposed for years to glycation, oxidation, and enzyme-mediated remodeling. Advanced glycation end products accumulate and create non-enzymatic cross-links that make collagen mechanically stiffer and harder to turn over. Lysyl oxidase family enzymes can further reinforce matrix structure under fibrotic conditions. This is not a cosmetic change. It directly alters tissue compliance and can trap organs in a low-plasticity state.
The second axis is compositional drift. Aging tissues often show more collagen deposition, thicker fibrils, basement-membrane alteration, proteoglycan changes, and a shift toward profibrotic architecture. In heart and lung this contributes to impaired compliance and fibrosis. In skeletal muscle it increases passive stiffness at the bundle level even when the muscle fibers themselves do not explain the full mechanical change. In vasculature it contributes to arterial stiffness, pulse-wave changes, endothelial dysfunction, and eventually a less forgiving hemodynamic system.
The third axis is signaling drift. Matrix proteins bind cytokines, growth factors, and cell receptors. Once the matrix changes, the signaling environment changes with it. A stiffer matrix can amplify fibroblast activation, reinforce myofibroblast states, and alter angiogenic behavior. Senescent cells worsen this problem by secreting proteases, inflammatory factors, and profibrotic cues that remodel the matrix further. The relationship therefore runs both ways: senescence damages matrix quality, and aged matrix sustains senescence-like dysfunction.
Why Cell Repair Alone Often Hits A Structural Ceiling
The easiest way to see the problem is to ask what happens after the cell-level intervention succeeds technically. Suppose stem-cell function improves, or a regenerative therapy delivers new cells, or partial reprogramming resets some transcriptional state. Those cells still have to operate inside a physical niche. If the surrounding matrix is stiff, inflammatory, poorly vascularized, and mechanically distorted, the new cells inherit bad instructions from the tissue context. They may engraft poorly, differentiate badly, or revert toward dysfunction.
This is why extracellular matrix aging belongs beside Stem Cell Exhaustion and Replenishment Strategies rather than below it. Stem-cell exhaustion is partly a cell-intrinsic problem, but the stem-cell niche is also matrix-dependent. A repaired cell inside a degraded scaffold is not the same as a repaired tissue. The same logic applies to senescence programs. Clearance of senescent cells may lower one source of inflammatory signaling, but if fibrosis and cross-link burden remain high, the tissue does not automatically return to youthful mechanics.
Inference, not settled fact: matrix damage may be one reason some rejuvenation platforms show cleaner biomarker or histology improvements than organ-level functional gains. This is a coherent inference because matrix mechanics influence force transmission, vascular behavior, and regenerative signaling across organs. It remains an inference because human interventional studies that isolate matrix restoration as the limiting variable are still sparse.

Where The Best Evidence Sits Today
The evidence for matrix aging as a real biological problem is strongest in descriptive and mechanistic domains. Human and animal studies repeatedly show increased stiffness, collagen accumulation, cross-linking, fibrosis, and altered mechanical behavior with age. Cardiac-aging literature documents collagen remodeling and fibrotic change. Skeletal-muscle work shows increased ECM contribution to passive stiffness in aged muscle. Angiogenesis research shows that an aged matrix impairs vessel sprouting and endothelial behavior. Reviews of lung fibrosis and senescence also show how age-modified ECM feeds a profibrotic loop.
The evidence for broad matrix rejuvenation is much weaker. There is no clinically mature intervention that generally restores youthful ECM architecture across organs in humans. Cross-link breakers have a long history of interest, but translation has been narrower and less decisive than early excitement implied. Anti-fibrotic drugs can matter in disease-specific settings, yet disease control is not the same thing as whole-body matrix rejuvenation. Tissue engineering and decellularized scaffold work show that matrix quality matters enormously, but they mostly confirm the importance of the problem rather than solve aging system-wide.
This asymmetry is why the topic is often neglected. The field can describe matrix aging clearly, but it cannot yet offer a simple consumer-facing intervention. Cell stories sell more easily because they suggest precision and reset. Matrix biology is harder because it is structural, distributed, organ-specific, and tightly linked to long-lived proteins that do not turn over quickly.
Where The Hype Usually Fails
The first mistake is to treat fibrosis, stiffness, and cross-linking as late-stage consequences rather than active drivers. They are both. Once matrix quality degrades, it reshapes cell behavior upstream and downstream. Calling the matrix secondary often means the model is too intracellular.
The second mistake is to assume that one intervention can generalize across all matrix problems. Collagen cross-link burden, elastin fragmentation, basement-membrane thickening, calcification, and scar remodeling are related but not identical pathologies. A drug that softens one fibrotic pathway is not automatically a global matrix-repair platform.
The third mistake is to confuse visible cosmetic change with deep matrix restoration. Skin appearance, transient hydration shifts, or short-term biomarker movement do not establish durable reversal of long-lived extracellular damage. Serious matrix repair would need to show changes in tissue mechanics, architecture, function, and durability, not only superficial signal.
What A Serious Matrix-Rejuvenation Standard Would Require
- Organ-specific endpoints. Vascular compliance, lung function, diastolic mechanics, skin elasticity, or muscle passive stiffness should be matched to the tissue actually being targeted.
- Direct structural measurement. Programs should quantify collagen content, cross-link burden, matrix organization, or stiffness, not rely only on blood biomarkers.
- Cell-context accounting. Trials should ask whether the intervention changes fibroblast state, endothelial behavior, stem-cell niche quality, or immune infiltration, because matrix effects are rarely isolated.
- Durability. A temporary anti-inflammatory benefit is not the same as structural matrix repair. The field needs evidence that architecture and function stay improved after the initial intervention window.
- Combination logic. The strongest future programs may pair cell repair, anti-fibrotic control, and matrix remodeling rather than treat them as competing silos.
This framework is stricter than the usual longevity pitch because it treats tissue architecture as a first-order variable. A rejuvenation claim that says nothing about the extracellular environment is often incomplete on contact.
Known, Inferred, And Unknown
| Category | Assessment |
|---|---|
| Known | Aging alters extracellular matrix composition, stiffness, and signaling across multiple organs through fibrosis, cross-linking, and remodeling drift. |
| Known | Matrix aging changes cell behavior through mechanotransduction and niche effects, which can impair regeneration, angiogenesis, and organ compliance. |
| Known | Human and animal evidence for matrix deterioration is much stronger than current evidence for broad, durable matrix rejuvenation. |
| Inferred | Some cell-focused longevity interventions likely underperform because they operate inside aged scaffolds that continue to impose dysfunctional mechanical and signaling conditions. |
| Unknown | Which combination of cross-link reduction, anti-fibrotic control, scaffold remodeling, and regenerative support can produce durable multi-organ ECM repair in humans without unacceptable tradeoffs. |
The Practical Reading For 2026
The disciplined conclusion is not that the extracellular matrix is the one true aging target. It is that serious longevity strategy cannot ignore it. A large share of tissue decline is architectural. If matrix damage is left outside the intervention model, the result may be cleaner mechanistic stories than real-world function. That is why this topic belongs with Hallmarks of Aging Revisited: What Changed Since 2013 and Senomorphics vs Senolytics: Slowing vs Removing Damage. All three point toward the same lesson: aging is not only a set of damaged cells. It is a damaged system.
For LifeMeter readers, the rule is practical. When a longevity claim promises tissue restoration, ask what it does to the scaffold, the niche, and the mechanics of the organ, not only to one molecular pathway. If it cannot answer that question, it may still be interesting science. It is not yet a complete repair strategy.
Further Reading Inside The Site
This article connects directly to Stem Cell Exhaustion and Replenishment Strategies, Hallmarks of Aging Revisited: What Changed Since 2013, and Senomorphics vs Senolytics: Slowing vs Removing Damage. Together they show why structural tissue context matters as much as cell state when longevity claims move toward translation.
Source List
Stearns-Reider KM, D'Amore A, Beezhold K, et al. Intrinsic stiffness of extracellular matrix increases with age in skeletal muscles of mice. Journal of Applied Physiology. 2015.
Wood LK, Kayupov E, Gumucio JP, et al. Alterations of Extracellular Matrix Mechanical Properties Contribute to Age-Related Functional Impairment of Human Skeletal Muscles. International Journal of Molecular Sciences. 2020.
Horn MA, Trafford AW. Aging and the cardiac collagen matrix: Novel mediators of fibrotic remodelling. Journal of Molecular and Cellular Cardiology. 2016.
Clarke SA, Richardson WJ, Holmes JW. The aged extracellular matrix and the profibrotic role of senescence-associated secretory phenotype. American Journal of Physiology. 2023.
Hwang J, Chambers E, Rustad KC, et al. Stiffening Matrix Induces Age-Mediated Microvascular Phenotype Through Increased Cell Contractility and Destabilization of Adherens Junctions. Advanced Science. 2022.
Spadaccio C, Rainer A, Trombetta M, et al. Impaired angiogenesis in ageing: the central role of the extracellular matrix. Clinical Science. 2023.
Lu N, Wang J, Li Y, et al. Extracellular matrix: new insights into its role in female reproductive aging and potential therapeutic strategies. npj Aging. 2026.
Translate this longevity claim into a capital-runway decision.
Life extension logic only matters if the balance sheet can carry it. Move into WealthMeter to compare assets, spending, and yield assumptions against the same long-horizon planning problem.