Stem Cell Exhaustion and Replenishment Strategies
Aging often shows up first as a slower recovery that people can feel before they can name it. Cuts close more slowly. Strength returns less fully after illness or injury. Tissues still contain stem cells, but the repair system around them has become biased, inflamed, stiff, and unreliable. The replenishment question is therefore not only how to add cells. It is how to restore a regenerative system that no longer gives those cells clear instructions.
The phrase entered the longevity canon because stem cell exhaustion was one of the original 2013 hallmarks of aging. The core observation still holds. Across blood, muscle, brain, intestine, and skin, tissue maintenance becomes less reliable with age because stem cells and their support environments lose regenerative quality. But the field has become more precise about what actually fails. Aging changes quiescence control, lineage bias, mitochondrial quality, epigenetic state, inflammatory tone, and extracellular-matrix architecture. The result is not one universal failure mode. It is a family of tissue-specific declines that share common systems logic. That sharper view matters because therapies aimed only at cell number can miss the older tissue conditions that make regeneration fail in the first place.
Core thesis: most credible replenishment strategies do not begin with wholesale stem-cell replacement. They begin by separating four different problems: depleted cell number, defective activation, biased differentiation, and damaged niche support. In many tissues, repairing the niche and recalibrating aged cells is more plausible in the near term than trying to repopulate an organ with newly delivered cells.
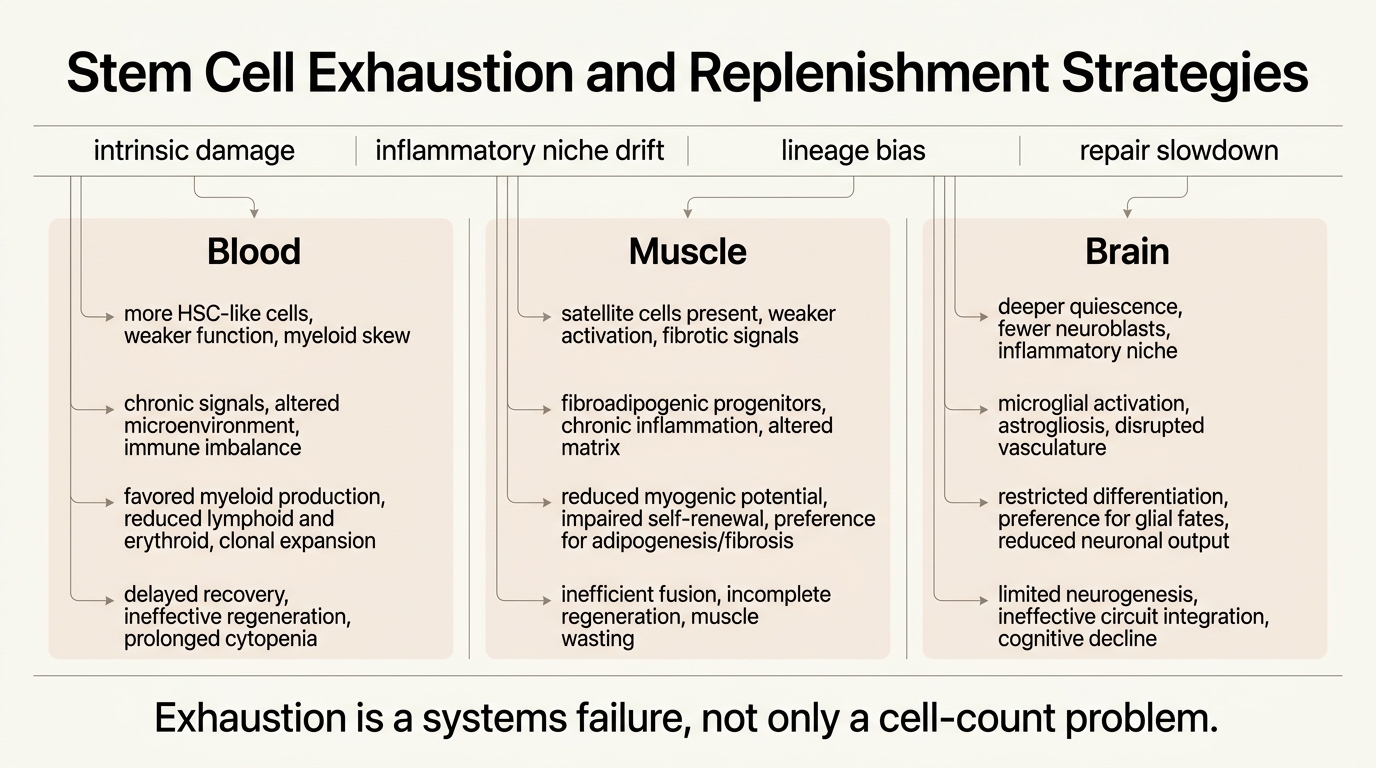
What Exhaustion Looks Like In Real Tissues
Hematopoietic stem cells show one of the clearest examples of why cell number alone misleads. Phenotypically defined HSC counts often increase with age, yet functional repopulating ability declines and lineage output skews toward myeloid cells. That combination helps explain why older immune systems become simultaneously inflamed, infection-prone, and malignancy-prone. More HSC-like cells do not mean a younger blood system if the pool is clonally biased and less regenerative.
Muscle tells a different story. Satellite cells often remain present into older age, but their activation and self-renewal logic degrades. Notch signaling weakens, TGF-beta and p38 stress signaling rise, and the local microenvironment becomes less supportive of repair. This is why muscle aging cannot be reduced to a simple shortage narrative. Aged muscle frequently contains cells that could contribute to regeneration if they were placed back inside a more permissive signaling context.
Neural stem cells present a third pattern. The old brain generally shows fewer activated stem cells and neuroblasts, deeper quiescence, and a more inflammatory niche. Even when some regenerative capacity remains, the brain is far less forgiving than blood or muscle because integration constraints are higher and damage from mis-specified cells is less acceptable. Replenishment in the brain therefore faces a stricter safety problem than the word regeneration tends to imply.
Why Replenishment Is Harder Than The Headline Suggests
Once exhaustion is understood as a systems defect, the limits of simplistic replenishment become obvious. Transplanted cells enter an old environment with altered cytokine signaling, fibrotic remodeling, vascular drift, immune dysfunction, and mechanical stiffening. If the niche remains aged, new cells may adopt aged behavior or fail to engraft at meaningful scale. This is one reason so much rejuvenation work now focuses on the stem-cell niche rather than on the stem cell in isolation.
The second problem is lineage control. Replenishment is only useful if the resulting cells generate the right differentiated progeny in the right proportions. A blood system skewed toward maladaptive myeloid output is not restored just because a transplant transiently increases cell number. The same logic applies in muscle, where proliferation without durable self-renewal or correct myogenic differentiation does not solve repair capacity. Regeneration requires architecture, not only proliferation.
The third problem is oncologic risk. Any strategy that increases proliferation, loosens epigenetic control, or pushes cells through partial dedifferentiation carries a surveillance burden. That burden is heavier in aging because aged tissues already accumulate mutations, clonal expansions, and immune dysfunction. The field can plausibly improve old regenerative systems. It cannot assume that every regeneration-boosting signal is automatically safe to repeat chronically in humans.
The Current Strategy Stack
The most serious replenishment programs fall into four buckets. The first is niche repair: lowering inflammatory noise, restoring extracellular-matrix quality, improving vascular support, and re-establishing signaling cues that let resident stem cells work again. Exercise, dietary interventions, and some pathway-targeted approaches fit here because they often improve regeneration indirectly by changing the environment around the cell.
The second bucket is intrinsic recalibration. This includes interventions aimed at pathways such as Notch, p38 MAPK, Cdc42, mitochondrial control, or epigenetic regulators that shift aged cells back toward younger functional states. Much of the excitement here comes from mouse and ex vivo systems showing that at least some aged stem cells are not irreversibly broken. They are mis-set. That distinction matters because a mis-set cell is more tractable than a depleted organ.
The third bucket is cell addition or replacement. Hematopoietic transplantation is the mature clinical proof that stem-cell replacement can work in a defined system, but it is also a reminder of how narrow the success conditions are: conditioning regimens, donor matching, graft-versus-host risk, and malignant-disease context all shape feasibility. Outside blood, large-scale replacement remains much harder. Delivery, engraftment, tissue integration, and long-run control remain unresolved in most aging applications.
The fourth bucket is partial reprogramming and hybrid regeneration strategies. These aim to restore youthful cell state without full loss of identity. The rationale is strong because epigenetic drift and loss of regenerative competence are real. The constraint is equally strong because the safety window is narrow and durable human evidence remains sparse. This is best understood as an upstream experimental layer, not as a near-term anti-aging service line.
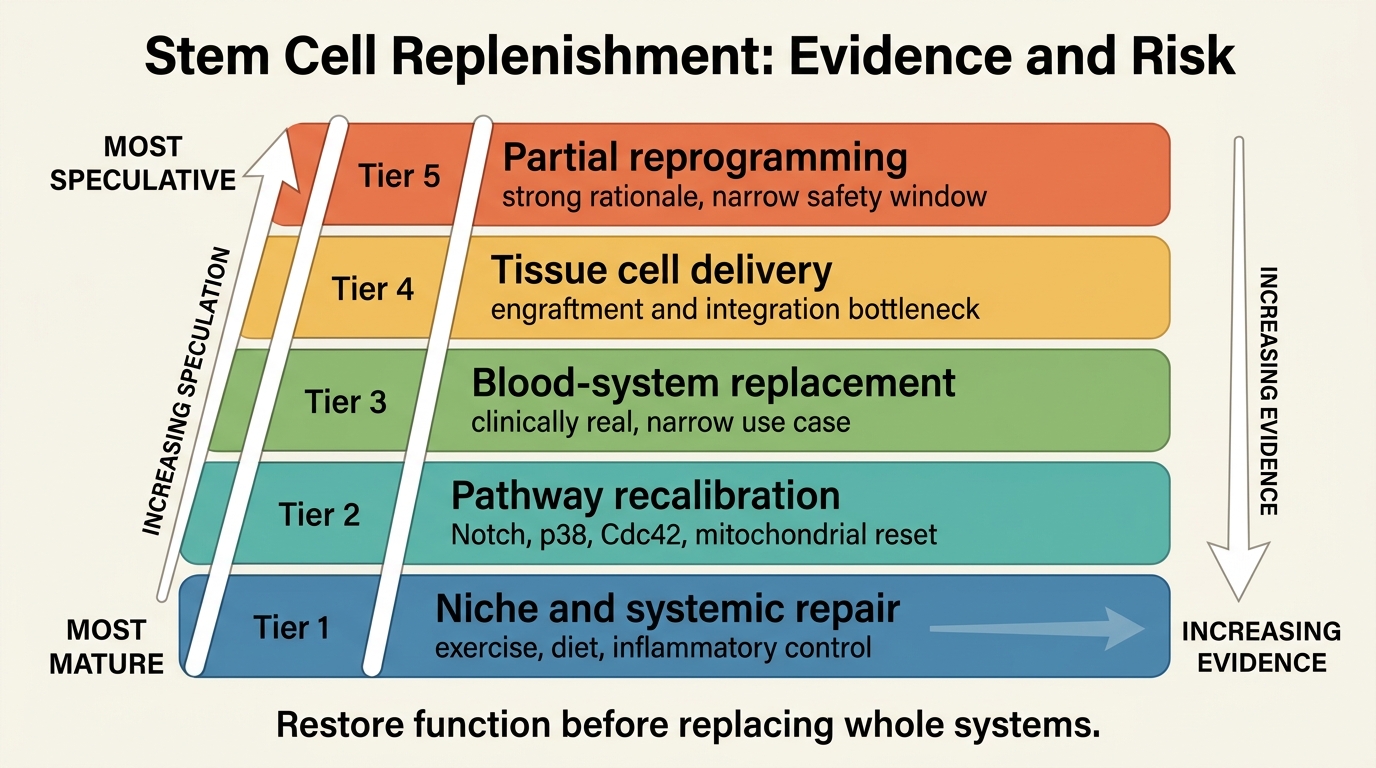
What The Evidence Supports Right Now
The strongest general conclusion is that aged stem cells are often constrained by both intrinsic and extrinsic aging, and the ratio varies by tissue. The broad Nature review of tissue stem-cell aging shows recurring themes across blood, brain, and muscle: reduced activation, altered progenitor output, inflammatory niche change, and systemic influences from diet, exercise, and circulating factors. That supports a systems model of exhaustion. It does not prove that one master intervention can reset all tissues at once.
The best-functioning replenishment logic today is therefore layered. Use niche and systemic interventions to improve the probability that resident cells behave well. Use targeted pathway repair where a tissue-specific bottleneck is reasonably clear. Reserve direct cell replacement for contexts where delivery and functional integration are already clinically tractable. This is less dramatic than the popular idea of injecting youth back into old organs. It is also more consistent with the evidence.
An important boundary follows from that. Replenishment is not the same as generalized rejuvenation. Restoring a stem-cell compartment can improve tissue repair, but it does not automatically solve inflammatory load, extracellular matrix damage, vascular aging, or organism-level frailty. Stem-cell exhaustion is a major node in aging biology. It is not the whole map.
Known, Inferred, And Unknown
| Category | Assessment |
|---|---|
| Known | Aging impairs stem-cell function across multiple tissues through both cell-intrinsic changes and niche deterioration. |
| Known | Hematopoietic stem cells can increase in number with age while losing functional quality and lineage balance. |
| Known | In muscle, at least some aged stem cells retain latent regenerative capacity that can improve when signaling conditions are recalibrated. |
| Inferred | Near-term progress is more likely to come from mixed strategies that restore resident function than from broad organ repopulation with exogenous cells. |
| Unknown | Which combinations can restore durable regeneration in humans without increasing malignancy, fibrosis, or maladaptive clonal expansion over time. |
What This Means For Longevity Work
Stem-cell exhaustion remains one of the most plausible leverage points in aging because it sits close to tissue maintenance itself. If stem-cell programs fail, repair slows, reserve shrinks, and downstream dysfunction spreads. But the translation lesson is restrictive. Credible regeneration programs will look less like generic stem-cell marketing and more like tissue-specific control systems with careful attention to inflammation, extracellular matrix, vascular support, lineage balance, and surveillance.
That is why the replenishment conversation has matured. Ten years ago, it was easier to frame old tissues as waiting for fresh cells. The stronger 2026 view is that old tissues are adaptive systems with degraded operating conditions. Some cells need replacement. Many need better instructions. Nearly all need a better environment. Replenishment is real. It is simply narrower, slower, and more engineering-heavy than the headline version suggests.
Further Reading Inside The Site
This article connects directly to Hallmarks of Aging Revisited: What Changed Since 2013, Partial Cellular Reprogramming: Reset Without Cancer Risk?, and Senolytics Moving into Clinical Translation. Those pieces map the adjacent questions of maintenance failure, reset biology, and damage clearance that shape whether replenishment can actually hold.
Source List
López-Otín C, Blasco MA, Partridge L, Serrano M, Kroemer G. The Hallmarks of Aging. Cell. 2013.
Brunet A, Goodell MA, Rando TA. Ageing and rejuvenation of tissue stem cells and their niches. Nature Reviews Molecular Cell Biology. 2023.
Rui C, Chan MKS, Skutella T. Stem Cell Therapies and Ageing: Unlocking the Potential of Regenerative Medicine. Subcellular Biochemistry. 2024.
Cosgrove BD, Gilbert PM, Porpiglia E, et al. Rejuvenation of the aged muscle stem cell population restores strength to injured aged muscles. Nature Medicine. 2014.
Zhang Y, Wang L, Zhao L, et al. Mechanisms and rejuvenation strategies for aged hematopoietic stem cells. Journal of Hematology and Oncology. 2020.
Li H, Geiger H. Cdc42 and aging of hematopoietic stem cells. Current Opinion in Hematology. 2014.
Translate this longevity claim into a capital-runway decision.
Life extension logic only matters if the balance sheet can carry it. Move into WealthMeter to compare assets, spending, and yield assumptions against the same long-horizon planning problem.