Yamanaka Factors in Vivo: Progress and Constraints
The field has cleared the novelty question. The serious issue in 2026 is no longer whether transient Yamanaka-factor expression can move aging-linked biology inside a living animal. It can. The question is whether the reset window can be engineered tightly enough to restore function without crossing into identity erosion, dysplasia, or a surveillance burden that makes the therapy unusable.
The public framing still misses the center of gravity. One story treats reprogramming as an imminent age-reset product. The other still treats it as inherently reckless embryology. Neither description matches the literature. Established fact: inducible expression of Oct4, Sox2, Klf4, and related partial combinations can alter age-linked molecular state and improve selected regenerative readouts in animal systems. Established fact: tissue targeting, expression control, dedifferentiation boundaries, and long-horizon safety remain unresolved. The field is neither empty nor deployable.
Core thesis: in vivo Yamanaka-factor work has produced real preclinical progress, especially in partial reprogramming and tissue-rescue models. The main barrier is no longer proof of reversibility. It is engineering a narrow reset window that improves aged tissue state without slipping into lineage drift, tumor risk, or an endpoint package too weak for clinical use.
What The Original Yamanaka Breakthrough Did And Did Not Prove
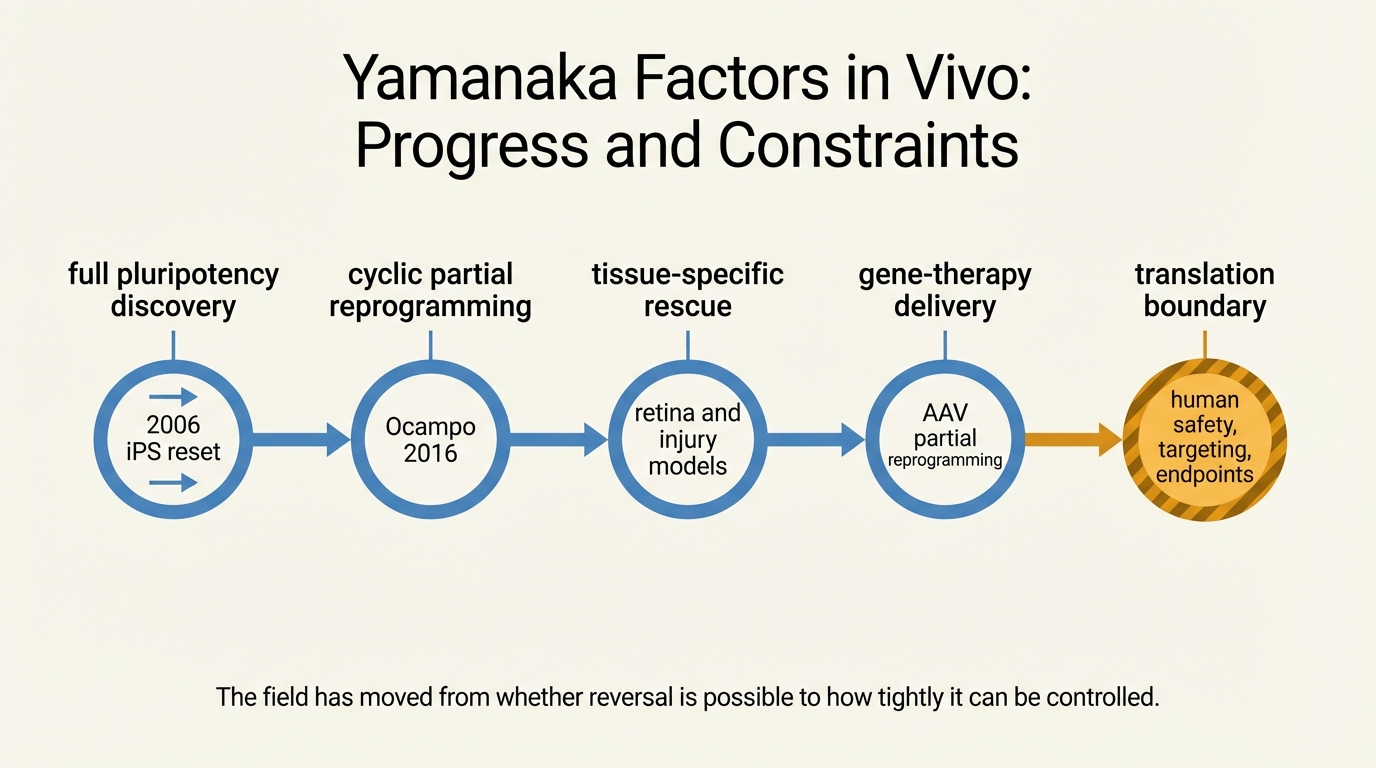
Takahashi and Yamanaka's 2006 work established that a small transcription-factor set could push differentiated cells back toward pluripotency. That result changed developmental biology and aging research because it showed that differentiated state is not irreversible. It did not show that an intact adult organism could be safely rejuvenated by reprogramming. Full reprogramming erases identity too far for most therapeutic purposes. In vivo, that creates an obvious hazard: a therapy intended to restore youthful function can instead destabilize tissue identity or produce teratoma-like growth.
The field therefore split quickly. One branch pursued induced pluripotent stem cells outside the body for replacement and disease modeling. The other, more relevant to longevity, asked whether brief or partial factor exposure could push old tissue state backward without crossing the point of no return. That second branch now contains most of the serious translational progress.
Where Real In Vivo Progress Appeared
Ocampo and colleagues provided the first strong animal proof that cyclic in vivo expression of OSKM could ameliorate age-associated phenotypes in a progeroid mouse model and improve regeneration after injury in older wild-type mice. That study did not solve the therapy problem, but it changed the burden of proof. After 2016, it was no longer reasonable to say that reprogramming inside an organism necessarily meant immediate catastrophic loss of tissue identity. Under tightly controlled schedules, beneficial partial reset appeared possible.
Progress became more concrete when the field shifted from whole-organism induction toward tissue-focused rescue. Lu and colleagues used OSK in retinal ganglion cells to restore youthful methylation patterns, promote axon regeneration, and improve vision-related outcomes in mouse eye models. That mattered because the intervention was local and the outputs were functional. This is a much stronger translational template than a broad claim about generalized rejuvenation without organ-specific leverage.
More recent gene-therapy work reinforced the same point. Cano Macip and colleagues reported lifespan extension and reversal of selected age-related changes in aged mice after AAV-mediated partial reprogramming. This does not establish a ready-made human therapy. It does establish that later-life intervention, viral delivery, and bounded factor expression can produce measurable biological effects in mammals. That is a meaningful shift from proof-of-principle novelty toward engineering reality.
Why Partial Reprogramming Is The Real Center Of Gravity
The phrase Yamanaka factors in vivo can mislead because it suggests one intervention. In practice the field has converged on a narrower logic: use transient, incomplete, and often tissue-limited expression to recover part of a youthful regulatory state while preserving lineage. That is very different from full pluripotent reversion. It is also why the strongest work emphasizes OSK rather than OSKM, bounded induction schedules, and disease contexts where regenerative upside can justify the residual uncertainty.
This is the conceptual advance. Aging biology has long treated epigenetic drift as one hallmark among many. Partial reprogramming treats that drift as something that might be manipulated inside living tissue rather than merely observed. If gene-expression state, chromatin configuration, and methylation architecture can be shifted toward a younger regime without erasing lineage, then part of age-linked dysfunction may be reversible. That proposition remains preclinical, but it is no longer loose speculation.
What remains uncertain is the width of the safe corridor. Partial reprogramming is attractive precisely because it tries to stop before dedifferentiation becomes pathological. That also means the effective window may be narrow, tissue dependent, and highly sensitive to delivery method, cell turnover, and injury context. A favorable result in retina or progeroid mice does not automatically generalize to liver, heart, brain, or systemic multi-organ use.
The Constraint Stack Is Now The Main Story
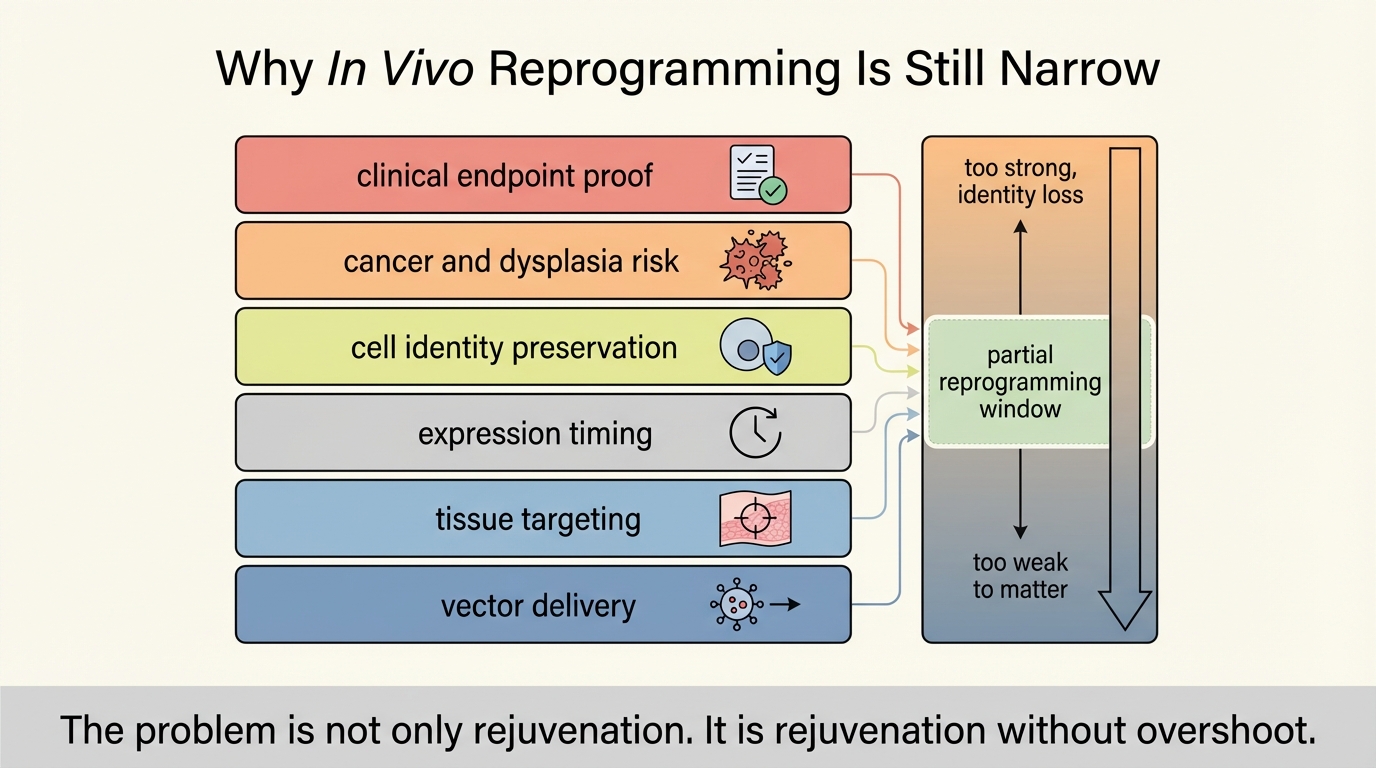
The first constraint is delivery. Distribution strong enough to reach relevant tissue is not the same as distribution safe enough to justify later repeat use. Viral delivery can work in defined contexts, but dose, tropism, immunogenicity, manufacturing, and retreatment feasibility all matter. Even if expression can be switched on, the field still needs reliable control over which cells receive the construct, how uniformly expression occurs, and how quickly it can be shut back down.
The second constraint is cell identity. Reprogramming is powerful because it perturbs the transcriptional architecture that makes a cell old. That same power can destabilize what makes the cell a neuron, hepatocyte, or cardiomyocyte. Therapeutic partial reprogramming only works if age-linked state improves faster than lineage fidelity erodes. That balance may differ sharply across tissues, disease contexts, and ages of intervention.
The third constraint is oncogenic and dysplastic risk. c-Myc is not incidental in the original cocktail. It is a potent growth driver with obvious cancer relevance. Even when protocols exclude c-Myc, the broader problem remains: interventions that loosen identity and proliferative controls can create delayed pathology that early rescue studies will not capture. The field has reduced the crude teratoma caricature. It has not removed the need for long-horizon safety evidence.
The fourth constraint is endpoint discipline. Many studies show favorable methylation or transcriptomic movement, but translation will depend on whether those shifts produce robust tissue function, disease modification, and acceptable safety margins. Biological-age marker movement is informative, but it is not itself a therapeutic endpoint. The same mismatch appears across longevity biotech: molecular plausibility often outruns clinically decisive outcomes.
What Is Established, What Is Inferred, What Is Still Unknown
Established fact: Yamanaka-factor biology demonstrated that cellular identity can be reset. Established fact: partial in vivo reprogramming can improve selected aging-related and regenerative outcomes in mice. Established fact: the strongest results depend on highly constrained experimental architectures rather than on any general whole-body rejuvenation protocol.
Reasoned inference: the first durable human use cases, if they emerge, are more likely to be tissue specific than organism wide. Ophthalmology, localized regenerative repair, or sharply defined organ dysfunction fit the evidence much better than a generalized aging indication. This remains an inference because no human efficacy pathway has yet validated it.
Unknown: whether partial reprogramming can be dosed repeatedly over years without cumulative instability. Unknown: whether the most useful factor set will remain close to classic OSK or move toward second-generation factor combinations and tighter control systems. Unknown: whether age-clock movement in treated tissue maps onto durable clinical benefit or only narrower transcriptional remodeling.
Why Human Translation Will Be Slower Than The Headlines Suggest
The mismatch between excitement and progress comes from conflating reversibility with deployability. Once a field shows that age-linked state can move backward in vivo, it is tempting to assume translation is mainly a matter of scale. It is not. Human therapy requires vector engineering, durable control systems, organ-specific toxicology, biodistribution mapping, manufacturing, and endpoint selection that can survive regulators and clinicians. A mouse-level success can be biologically profound and still represent only the beginning of the real therapeutic program.
This does not make the field empty hype. The preclinical signal is strong enough to justify serious investment and continued mechanistic work. It does mean the useful framing must narrow. Yamanaka factors in vivo are best understood as a platform-level attempt to manipulate epigenetic state, not as a single impending anti-aging product. Platform work advances through control problems, and those are exactly the problems still open.
Known, Inferred, And Unknown
| Category | Assessment |
|---|---|
| Known | Transient in vivo expression of Yamanaka-factor combinations can improve selected regeneration and aging-linked phenotypes in animal models. |
| Known | The strongest preclinical progress comes from partial, cyclic, or tissue-specific expression rather than unrestricted whole-body reprogramming. |
| Known | Delivery control, lineage preservation, long-horizon safety, and endpoint validity remain the central translational constraints. |
| Inferred | Early human translation is more likely to succeed in local or organ-specific indications than in generalized aging treatment. |
| Unknown | Whether repeated human use can deliver clinically meaningful rejuvenation without dysplasia, cancer risk, or progressive tissue-identity loss. |
The Practical Reading For 2026
The correct LifeMeter reading is neither dismissal nor product-level optimism. Yamanaka-factor work in vivo deserves to be treated as one of the most consequential reversibility programs in aging biology. It has already produced enough animal evidence to clear the threshold of intellectual seriousness. But it remains a control-limited technology. The hard problem is not proving that cells can move backward. The hard problem is telling them how far, where, and for how long without paying an unacceptable price.
That is why this topic sits downstream of Partial Cellular Reprogramming and alongside Epigenetic Reprogramming Enters Human Trials. The biology of reset is real. The therapeutic corridor is still narrow.
Further Reading Inside The Site
This article connects directly to Partial Cellular Reprogramming: Reset Without Cancer Risk?, Epigenetic Reprogramming Enters Human Trials, and Stem Cell Exhaustion and Replenishment Strategies. Together they separate biological reversibility from translational control.
Source List
Takahashi K, Yamanaka S. Induction of Pluripotent Stem Cells From Mouse Embryonic and Adult Fibroblast Cultures by Defined Factors. Cell. 2006.
Ocampo A, Reddy P, Martinez-Redondo P, et al. In Vivo Amelioration of Age-Associated Hallmarks by Partial Reprogramming. Cell. 2016.
Lu Y, Brommer B, Tian X, et al. Reprogramming to Recover Youthful Epigenetic Information and Restore Vision. Nature. 2020.
Cano Macip C, Hasan R, Hoznek V, et al. Gene Therapy-Mediated Partial Reprogramming Extends Lifespan and Reverses Age-Related Changes in Aged Mice. Cell Reprogramming. 2024.
Cho HE, Lee S, Seo JH, et al. In Vivo Reprogramming Using Yamanaka Factors in the CNS: A Scoping Review. Cells. 2024.
Translate this longevity claim into a capital-runway decision.
Life extension logic only matters if the balance sheet can carry it. Move into WealthMeter to compare assets, spending, and yield assumptions against the same long-horizon planning problem.