Network Biology of Aging: Identifying Master Regulators
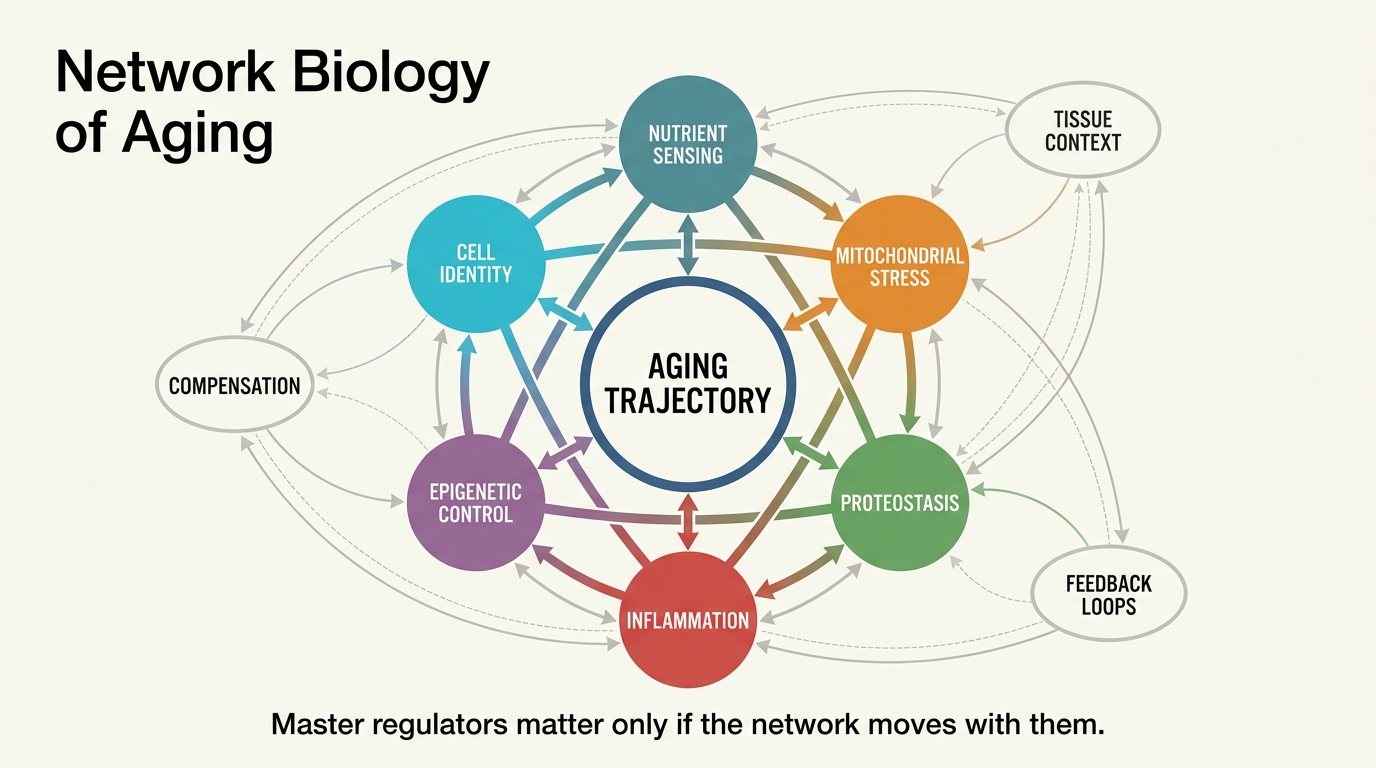
Aging does not unfold as one broken pathway ticking upward on a clean chart. It behaves more like a coupled control system in which stress signaling, metabolism, repair capacity, cell identity, and tissue environment constantly reshape one another. Network biology matters because it tries to model that coupling directly. It becomes misleading when one inferred hub is treated as if it were a universal rejuvenation switch.
The attraction of master regulators is obvious. If aging is a network problem, then perhaps a few central nodes explain most of the variance and offer the best intervention leverage. Sometimes that logic is productive. A transcription factor, signaling axis, or regulatory program can sit high enough in the system to coordinate broad downstream effects. The difficulty is that biological networks are layered, adaptive, and tissue-dependent. Centrality on one map does not automatically mean safe or sufficient therapeutic leverage in a living organism.
Core thesis: network biology is one of the strongest frameworks for understanding aging because it treats decline as an interacting system rather than a set of isolated lesions. The established fact is that aging-related regulation is distributed across coupled transcriptional, metabolic, inflammatory, and tissue-specific networks. The inference, often overstated, is that identifying a central regulator means one target can reset the system broadly enough to count as rejuvenation.
Why Aging Needs A Network Frame
Most familiar aging categories already imply network logic. Mitochondrial decline affects redox signaling, proteostasis, inflammation, and stem-cell function. Senescent cells alter neighboring tissue through secreted factors. Epigenetic drift changes which repair programs are even available to a cell. None of those processes remains contained within a single compartment for long. That is why pathway-by-pathway storytelling often compresses the biology too aggressively.
Network biology does better because it asks how regulatory relationships are organized, which nodes influence multiple downstream programs, and where feedback loops amplify or dampen damage. In practice, that can mean gene-regulatory networks, protein interaction maps, cell-cell communication models, or multi-omics integration layers that infer which control modules shift with age. The model class is broader than one technique. The common idea is that influence matters more than raw list-making.
This shift is not cosmetic. It changes what counts as evidence. A long gene list is descriptive. A regulator that consistently organizes multiple age-linked states across tissues, cell types, or interventions is more mechanistically interesting. That is the real value of the network frame.
What The Strongest Papers Actually Show
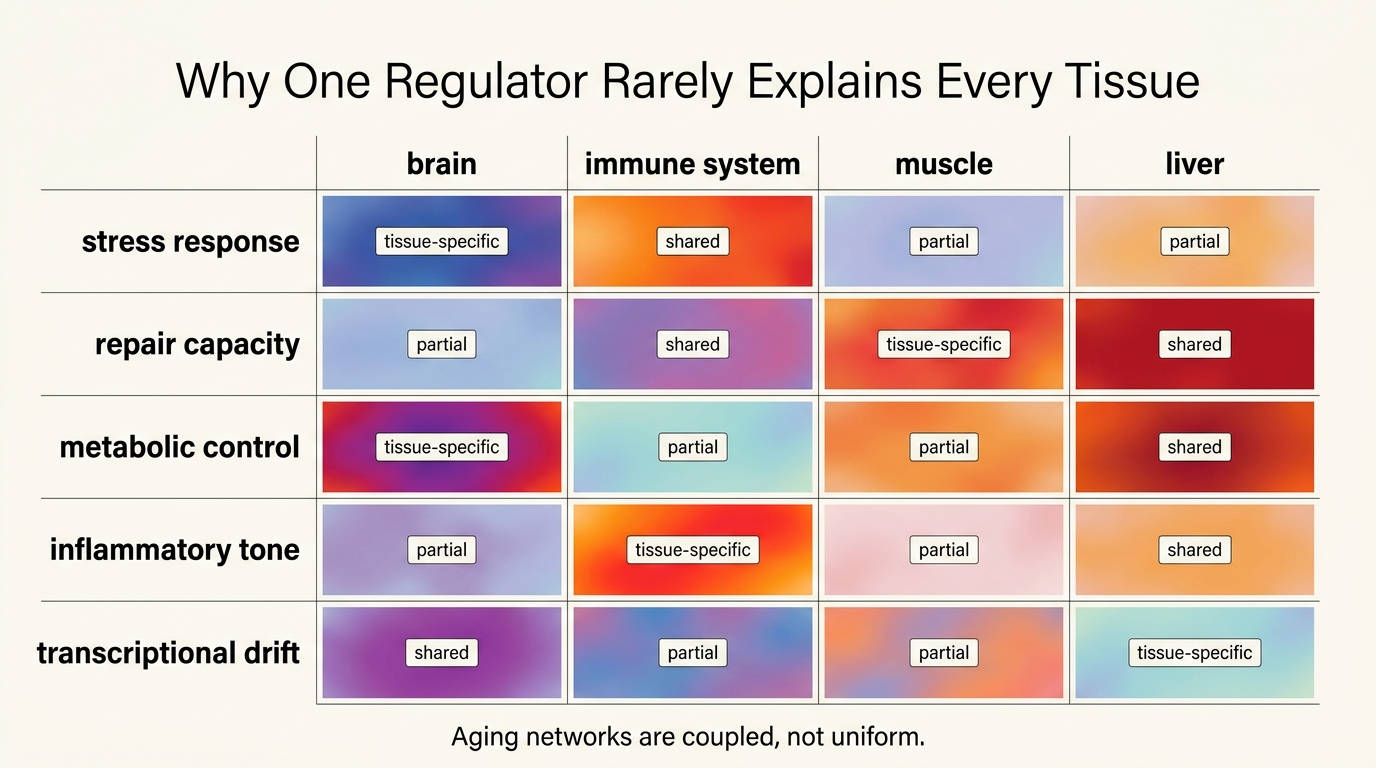
Recent systems work has improved this picture in two ways. First, large-scale organism and tissue maps show that aging signatures are not uniform across the body. The Stanford multi-omics atlas in Nature found organ-specific temporal patterns rather than one synchronized decay curve. That matters because a central regulator in liver or immune cells may not govern the same way in brain or muscle.
Second, newer regulatory-network methods are beginning to map age-associated control structure rather than only endpoint markers. The 2024 Cell study on organism-scale gene-regulatory networks used population asynchrony to infer regulatory organization across aging trajectories. The important implication is not that one molecule has already been crowned the aging master key. It is that network-level inference is becoming precise enough to distinguish broad control modules from downstream noise.
Single-cell work points in the same direction. Nature Aging analyses of age-associated transcription-factor activity show that regulatory drift can be cell-type specific, meaning the same organism contains multiple local aging-control problems at once. That is one reason simple universal-reset claims usually fail under closer inspection.
What Counts As A Master Regulator
The phrase is often used too loosely. A true master regulator should meet a higher bar than mere correlation. It should sit upstream of multiple downstream processes, show reproducible control across relevant contexts, and move a meaningful fraction of system state when perturbed. Even then, the intervention question remains open. A node can be central yet pharmacologically inaccessible, unsafe to manipulate chronically, or too entangled with normal function to serve as a clean therapeutic lever.
This distinction matters because aging research often promotes high-level categories such as mTOR, AMPK, FOXO, NF-kB, or chromatin-state regulators as if centrality alone settled the therapeutic case. Those nodes do matter. The harder question is whether moving them in adult humans will produce durable functional benefit across tissues without intolerable tradeoffs. Network logic sharpens that question. It does not erase it.
Readers should also distinguish between a master regulator of one local program and a master regulator of organism-wide aging. Those are not interchangeable claims. The field has much more evidence for the first than the second.
Why Tissue Context Keeps Winning
Aging is local before it is global. Immune aging, glial aging, muscle aging, and stromal aging each involve different constraints, even when some upstream motifs overlap. Tissue architecture, turnover rate, repair reserve, and exposure to metabolic or inflammatory stress all shape which parts of the network matter most. That is why the same intervention can look elegant in one tissue and marginal in another.
This is also why the topic connects directly to tissue-specific senescence, digital twins for aging, and multi-omics integration. The future likely belongs to layered models that identify both shared control logic and tissue-bound exceptions. A universal network map with no local context is too coarse to guide intervention well.
What Network Biology Is Good For Right Now
- Target ranking. It helps prioritize which regulators are upstream enough to deserve experimental attention.
- Combination logic. It can reveal when one pathway intervention is unlikely to move the system without a second supportive change.
- Biomarker interpretation. It is better at separating downstream movement from real control-shift candidates.
- Tissue comparison. It clarifies whether one signal is shared across organs or mostly local.
- Failure analysis. It helps explain why clean single-pathway interventions often underperform in aged organisms.
What It Is Not Good Enough For Yet
It is not yet a license to talk as if the field has already isolated the switch that governs whole-body aging. Many network models still depend on inference quality, dataset bias, incomplete temporal coverage, and assumptions about causality that remain only partly tested. A regulator that looks central in transcriptomic or accessibility data may still be one layer removed from the true intervention bottleneck.
There is also a translation gap. Network maps are often strongest at telling us where to look next, not at guaranteeing that a tractable medicine exists at the identified node. For LifeMeter readers, that means systems biology should increase confidence in disciplined mechanistic prioritization, not in near-term claims of broad age reset.
Known, Inferred, And Unknown
| Category | Assessment |
|---|---|
| Known | Aging signatures differ across organs and cell types, and system-level multi-omics studies consistently show non-uniform temporal behavior across tissues. |
| Known | Gene-regulatory and transcription-factor analyses can identify age-associated control modules that are more informative than isolated marker lists. |
| Known | Centrality in a network model does not by itself prove safe, sufficient, or organism-wide therapeutic control. |
| Inferred | The most useful near-term role for network biology is target ranking and combination design rather than single-node rejuvenation claims. |
| Inferred | Many apparent master regulators will prove tissue-weighted rather than globally dominant once perturbation data accumulates. |
| Unknown | Which inferred control nodes will remain causal under longitudinal human perturbation, and which will collapse into context-dependent correlates once translated beyond model systems. |
Practical Reading For LifeMeter
The network frame improves thinking because it disciplines the field against one-pathway fantasy. It reminds readers that aging is a control problem with cross-coupled subsystems, feedback loops, and tissue-specific expression. That is a more realistic map than any one-molecule doctrine.
The same realism should also make readers more selective. A credible master-regulator claim should show cross-context influence, perturbation relevance, and some reason to believe the intervention can move function rather than only signatures. Until then, network biology is best viewed as a high-value discovery engine and interpretation layer, not yet as proof that one knob can reset the organism.
Source List
Schaum N, Lehallier B, Hahn O, et al. Ageing hallmarks exhibit organ-specific temporal signatures. Nature. 2020.
Eder M, Martin OMF, Oswal N, et al. Systematic mapping of organism-scale gene-regulatory networks in aging using population asynchrony. Cell. 2024.
Hao Y, Hao S, Andersen-Nissen E, et al. Inference of age-associated transcription factor regulatory activity changes in single cells. Nature Aging. 2022.
Avila PU, Padvitski T, Leote AC, et al. Gene regulatory networks in disease and ageing. Nature Reviews Nephrology. 2024.
Translate this longevity claim into a capital-runway decision.
Life extension logic only matters if the balance sheet can carry it. Move into WealthMeter to compare assets, spending, and yield assumptions against the same long-horizon planning problem.