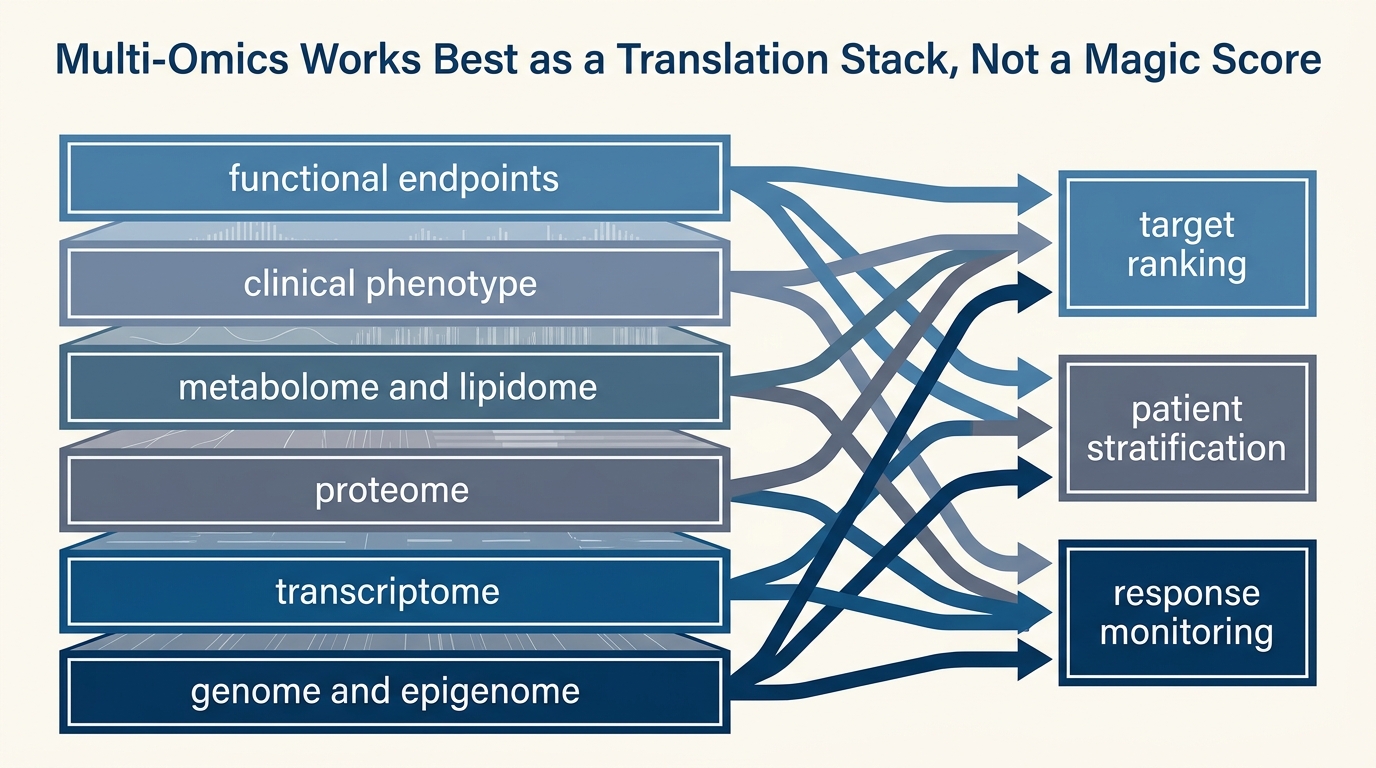
Multi-Omics Integration: From Data to Intervention
Longevity medicine has reached the point where the bottleneck is no longer raw biomarker generation. Labs can measure methylation, proteins, metabolites, inflammatory cytokines, microbiome shifts, and clinical chemistry at scale. The harder problem is translation. A large molecular panel can describe a body in high resolution and still fail to answer the practical question that matters most: what should be changed, in whom, and with what level of confidence that the change will improve a real health outcome rather than only move an intermediate signal.
That distinction matters because multi-omics is often marketed as if measurement complexity itself creates intervention clarity. The current evidence does not support that shortcut. What the field does support is narrower and more useful. Multi-omics can reveal aging patterns that single-layer assays miss, improve stratification, identify candidate pathways worth testing, and monitor how an intervention perturbs a system. What it still cannot do reliably is convert correlation-rich molecular signatures into a clinically validated anti-aging therapy without a separate causal bridge.
Core thesis: multi-omics is strongest today as a discovery, stratification, and monitoring stack. It becomes weak when its users treat dense molecular correlation as a substitute for mechanism, tissue specificity, or clinical endpoint validation.
Why the field moved beyond single-layer biomarkers
The aging-biomarker literature now treats single readouts with more caution than the public narrative usually does. The 2023 Cell consensus on biomarkers of aging argued that useful aging biomarkers should do more than correlate with chronological age. They should ideally predict functional decline or mortality better than chronological age and show responsiveness to interventions targeting aging. That standard immediately exposes the limitation of one-dimensional measurement. A methylation clock can be predictive without explaining which tissue is driving the signal. A plasma protein signature can correlate with mortality while remaining too aggregated to choose a specific intervention. A clinical panel can reveal metabolic stress but miss upstream regulatory structure.
Multi-omics became attractive because aging is not executed on one molecular layer. Genomic instability, altered transcriptional programs, proteostatic stress, metabolic rewiring, inflammation, immune drift, and microbiome-mediated signaling all interact. A single marker can be useful. It is rarely sufficient. Integration is the attempt to move from isolated readouts toward a systems view in which several layers help constrain one another.
What integration already revealed that linear aging models missed
A strong recent example came from the Stanford-led Nature Aging study published in August 2024. The authors followed 108 adults aged 25 to 75 years in a longitudinal cohort and collected ten categories of omics and clinical data, including transcriptomics, proteomics, metabolomics, cytokines, lipidomics, laboratory measures, and several microbiome layers. Their central finding was not only that aging is measurable at multiple molecular levels. It was that many molecular changes are nonlinear, with substantial dysregulation clustering around about age 44 and again around about age 60.
That result matters for intervention design. If biological transitions are clustered rather than smoothly linear, then the best intervention windows may also be clustered. A prevention program aimed at a 42-year-old with early cardiometabolic drift may not look like the program for a 61-year-old whose inflammatory and renal signals are changing together. Single-layer models often blur these transitions into one continuous slope. Multi-omics can separate them into distinct system shifts.
The implication here is reasoned inference, not proof. The study identified nonlinear transitions and pathway shifts. It did not prove that intervening at those transition points changes long-run outcomes. Still, it offers a more realistic map for where targeted trials should be designed.
Where the evidence is strongest right now
The strongest current use case is not “multi-omics found the longevity drug.” It is “multi-omics improves measurement of biological state in ways that can support target ranking and response monitoring.” Three lines of evidence illustrate that narrower claim.
First, molecular aging clocks are becoming more predictive across populations. The August 2024 Nature Medicine proteomic aging clock study showed that a plasma-protein-based clock predicted mortality and risk of common age-related diseases across diverse cohorts. That is a meaningful advance because proteins often sit closer to active physiology than upstream nucleic signals alone. It does not mean the proteomic clock by itself selects therapy. It means the clock captures clinically relevant age-associated state.
Second, clock construction is becoming more interpretable. In June 2024, Nature Aging published a principal-component-based clinical aging clock that identified signatures of metabolic dysregulation, cardiac and renal dysfunction, and inflammation associated with unsuccessful aging. The important point was not merely prediction. The authors also showed that those processes can be impacted by established drug interventions. That is closer to the translational ideal because the clock dimensions are linked to pathways that clinicians already understand.
Third, intervention studies can now register tissue-level response patterns rather than only surface biomarkers. A 2024 Nature Communications secondary analysis of the GOTO lifestyle-intervention study integrated transcriptomic responses across tissues in older adults and demonstrated coordinated molecular response to a combined lifestyle intervention. Lifestyle interventions are not exotic geroscience, but they are useful proof that an intervention can perturb a multi-tissue molecular state in structured rather than random ways.
| What is established | What is inferred | What is still unknown |
|---|---|---|
| Multi-omics captures age-associated molecular state more comprehensively than single-layer panels. | Layered measurements can improve identification of transition windows and intervention subtypes. | Whether acting on multi-omics signatures alone improves long-term healthspan in humans. |
| Some clocks and integrated signatures predict mortality, disease risk, or unsuccessful aging better than age alone. | Signals that converge across layers are more likely to be mechanistically relevant than isolated biomarkers. | Which integrated signatures are robust enough to serve as surrogate endpoints for anti-aging drug approval. |
| Interventions such as lifestyle change can produce coordinated multi-tissue molecular responses. | Integrated monitoring can probably improve dose finding and early response assessment in trials. | How often short-term molecular improvement translates into durable functional benefit. |
Why the translation gap remains real
The gap persists because molecular integration does not remove the classical problems of causal inference. It reorganizes them. A strong age-associated signal may be upstream damage, compensatory adaptation, tissue leakage, exposure history, medication effect, or simple cell-composition shift. The more data layers a model contains, the more convincing a correlation can look without necessarily becoming more causal.
The 2025 Nature Reviews Genetics review on epigenetic clocks makes this point directly. Epigenetic clocks are powerful machine-learning tools, but their interpretation is limited by computational and statistical challenges including cell-type heterogeneity. A bulk blood methylation signal can change because the biology of a cell changed, because the relative abundance of cell populations changed, or because both happened at once. Multi-omics reduces some ambiguity by comparing layers, but it does not eliminate it.
Tissue specificity is the second major constraint. Blood is convenient, repeatable, and clinically scalable. Aging is organ-specific in many of its practical failures. A composite plasma signature may tell you that biological aging burden is rising while remaining too coarse to choose between an intervention aimed at immune regulation, mitochondrial dysfunction, renal stress, or neurovascular decline. The integrated signal is real. The intervention pathway can still be underspecified.
Temporal mismatch is the third constraint. Molecular layers operate on different clocks. Transcripts can change in hours. proteins and metabolites may show medium-horizon adaptation. Clinical endpoints such as frailty, disability, or survival move on far slower timescales. An intervention can therefore produce a molecular response that looks impressive long before anyone knows whether it represents true repair, temporary compensation, or measurement artifact.
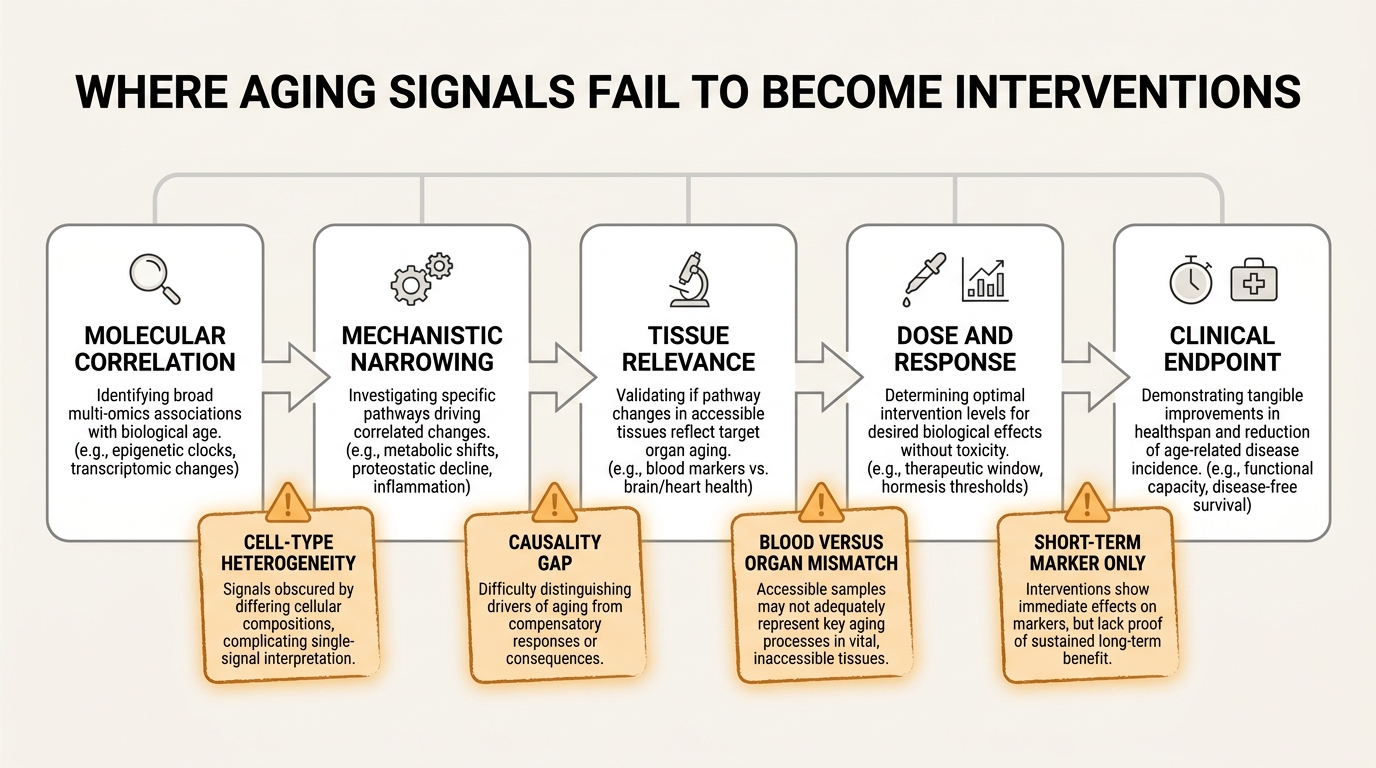
What a credible intervention pipeline should look like
The most defensible pipeline is staged. Multi-omics should first identify a reproducible signal across cohorts. Next, the signal should be decomposed into pathways that are biologically interpretable and tissue-relevant. Then an intervention should be tested against that pathway with short-horizon pharmacodynamic markers and a separate plan for longer-horizon functional endpoints. Only after those layers align should the field speak confidently about intervention relevance.
- Discovery: identify convergent multi-layer signatures that persist after adjustment for obvious confounders.
- Mechanistic narrowing: determine whether the integrated signature reflects likely driver biology, downstream damage, or compensatory response.
- Patient stratification: use the integrated profile to define which subgroup is actually being targeted.
- Pharmacodynamic monitoring: confirm that the intervention moves the intended pathway, not only a generic wellness marker.
- Clinical validation: show that the molecular change connects to function, disease incidence, or another endpoint that matters outside the assay environment.
That pipeline is less glamorous than claims about a universal biological age score, but it is how the field becomes clinically serious. Multi-omics should not be treated as a replacement for trials. It should be treated as an instrument panel that can make better trials possible.
What readers should trust and what they should reject
Readers should trust multi-omics most when it is used to make a narrow claim with explicit boundaries. Good examples include identifying a reproducible inflammatory aging signature, stratifying participants who appear metabolically older than their chronological age would suggest, or showing that a drug candidate perturbs a defined pathway across several layers of measurement. Those are meaningful advances.
Readers should reject the stronger and more common leap: an integrated age score moved, therefore the therapy is anti-aging in a clinically durable sense. That conclusion still requires more than dense molecular correlation. It requires evidence that the shift is mechanistically coherent, repeatable, and connected to outcomes that are not reducible to the assay itself.
The field is moving in the right direction. Multi-omics is not hype in the trivial sense. It is genuinely changing how aging biology is measured. The discipline required now is not more enthusiasm for data density. It is stricter separation between molecular description, mechanistic inference, and validated intervention.
Primary Sources
Moqri M, Herzog C, Poganik JR, et al. Biomarkers of Aging for the Identification and Evaluation of Longevity Interventions. Cell. 2023.
Shen X, Wang C, Zhou X, et al. Nonlinear dynamics of multi-omics profiles during human aging. Nature Aging. 2024.
Fong S, Pabis K, Latumalea D, et al. Principal component-based clinical aging clocks identify signatures of healthy aging and targets for clinical intervention. Nature Aging. 2024.
Argentieri MA, Xiao S, Bennett D, et al. Proteomic aging clock predicts mortality and risk of common age-related diseases in diverse populations. Nature Medicine. 2024.
Bogaards FA, Gehrmann T, Beekman M, et al. Secondary integrated analysis of multi-tissue transcriptomic responses to a combined lifestyle intervention in older adults from the GOTO nonrandomized trial. Nature Communications. 2024.
Teschendorff AE, Horvath S. Epigenetic ageing clocks: statistical methods and emerging computational challenges. Nature Reviews Genetics. 2025.
Translate this longevity claim into a capital-runway decision.
Life extension logic only matters if the balance sheet can carry it. Move into WealthMeter to compare assets, spending, and yield assumptions against the same long-horizon planning problem.