Reversible Aging Signatures in Animal Models
Aging used to be described as a one-way accumulation problem. Damage rises, repair capacity falls, and tissues slide downhill until the slope becomes clinically visible. Animal work no longer supports such a simple one-direction picture. In several model systems, some age-linked signatures can be pushed backward. The hard part is defining what “backward” actually means.
A reversed methylation clock is not the same thing as regenerated tissue. Better muscle repair is not the same thing as durable cancer-safe rejuvenation. Improved old-mouse physiology after blood exchange or partial reprogramming is not the same thing as proving that aging as a whole has been turned around. The field is now strong enough that dismissal is unserious, but still incomplete enough that sweeping victory language is wrong.
Core thesis: animal models do show that some age-linked states are reversible. The strongest evidence sits in specific tissues, specific signatures, and bounded interventions such as partial reprogramming, regenerative injury models, and prolonged systemic-environment manipulations. What remains unproved is whole-organism reversal that is durable, broadly safe, and functionally integrated across tissues.
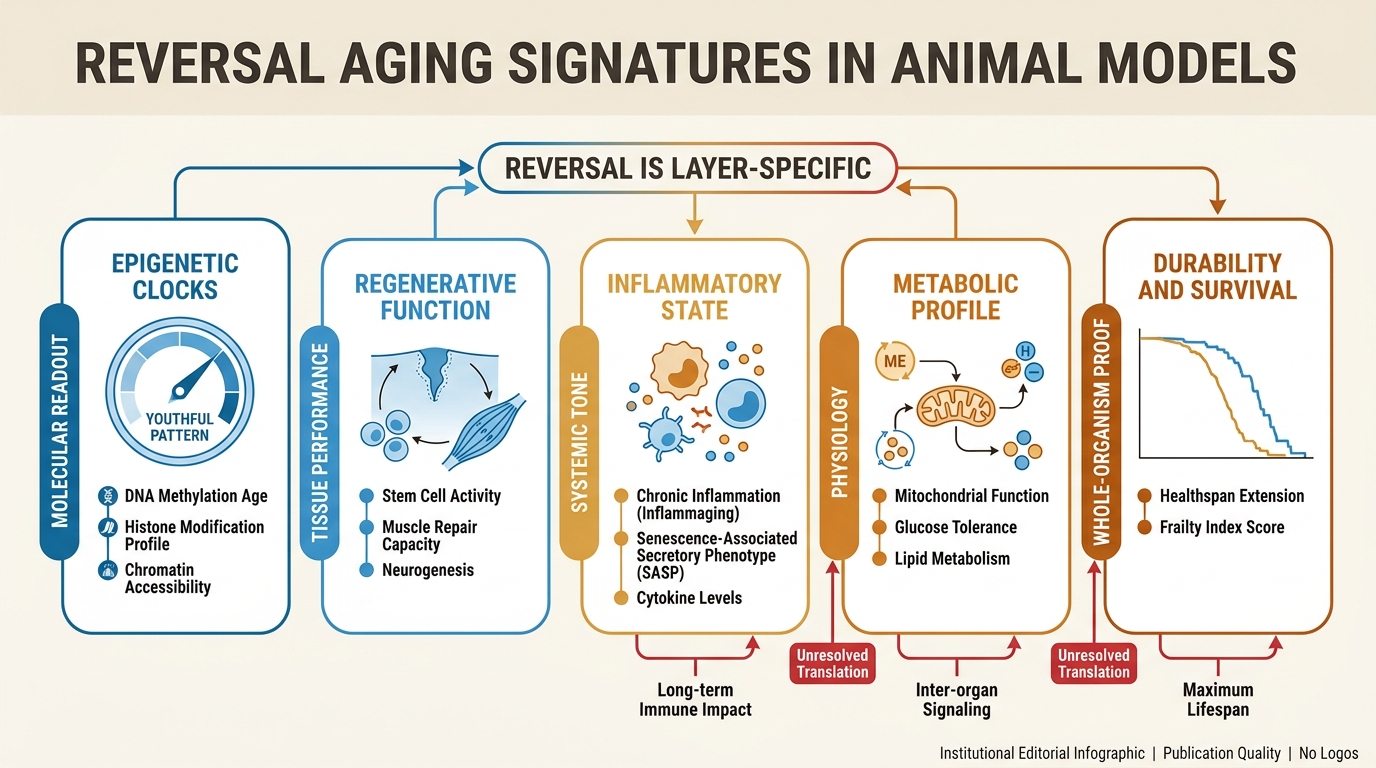
What Counts as a Reversal Signal
The cleanest starting point is to separate age-linked readouts into layers. Some are molecular summaries, such as methylation signatures or transcriptomic profiles. Some are functional, such as regeneration after injury, muscle performance, cognition, or frailty scores. Some are structural, such as fibrosis burden, tissue architecture, or stem-cell composition. Some are organism-level outcomes such as healthspan or lifespan. A serious reversal claim should say explicitly which layer moved.
This matters because the layers do not always travel together. A methylation age estimate can improve without proving that every relevant tissue regained youthful repair capacity. A local regeneration result can be striking without showing that the broader aging process was reset. The right reading is therefore layered rather than absolute. Animal models establish that at least part of aging biology remains plastic. They do not yet establish that all major age-related failure modes are jointly reversible by one operating principle.
That is also why methylation drift and biological age clocks have to be handled carefully. They are useful because they can register movement faster than lifespan endpoints, but they can also tempt the field into treating a proxy as a verdict.
Where the Strongest Animal Evidence Sits Today
The most credible reversal literature clusters into three buckets. The first is partial reprogramming, where transient expression of reprogramming factors shifts molecular age markers and, in some models, improves tissue function. The second is regenerative rescue in injured or aged tissues, especially in optic-nerve and retinal-ganglion-cell work, where mature neurons regained more youthful repair behavior after targeted reprogramming. The third is systemic-environment work, including heterochronic parabiosis and related exchange paradigms, where old animals exposed to younger systemic context showed younger molecular signatures and in some studies better physiological outcomes.
None of those buckets is trivial. They collectively defeat the old claim that age-linked tissue states are fixed once adulthood is reached. They show instead that at least some hallmarks of aged state are maintained dynamically and can be moved when the regulatory environment changes enough. That is the real breakthrough.
They also differ in what they prove. Partial reprogramming is strongest as a cell-state intervention and weakest where long-run control, tumor risk, and cross-tissue governance still dominate. Systemic exchange models are strongest as evidence that circulating factors and systemic signaling matter more than once assumed, but weakest as direct therapeutic templates. Tissue regeneration studies are strongest where the endpoint is sharp and local, but weaker as whole-body inference.
Why Reprogramming Results Matter More Than The Headlines Around Them
Reprogramming work matters because it is the clearest evidence that age-linked epigenetic state is not merely passive residue. If transient factor expression can move cells toward a younger functional profile, then at least part of biological aging is being stabilized by reversible regulatory architecture. That does not mean the stronger “information theory solves aging” frame is already proved. It means the plasticity question has moved from speculation to constrained evidence.
The critical constraint is control. As argued in Yamanaka Factors in Vivo: Progress and Constraints, the bottleneck is no longer whether reset signal exists. It is whether the reset can be bounded tightly enough to improve tissue state without eroding cell identity, triggering dedifferentiation, or drifting into oncogenic risk. A field can be biologically real and still therapeutically immature.
This is where animal models are most informative. They show that timing, tissue, delivery method, and endpoint choice matter enormously. Reversal is not a uniform bath that washes over the whole organism. It is a controlled perturbation whose benefit depends on operating window.
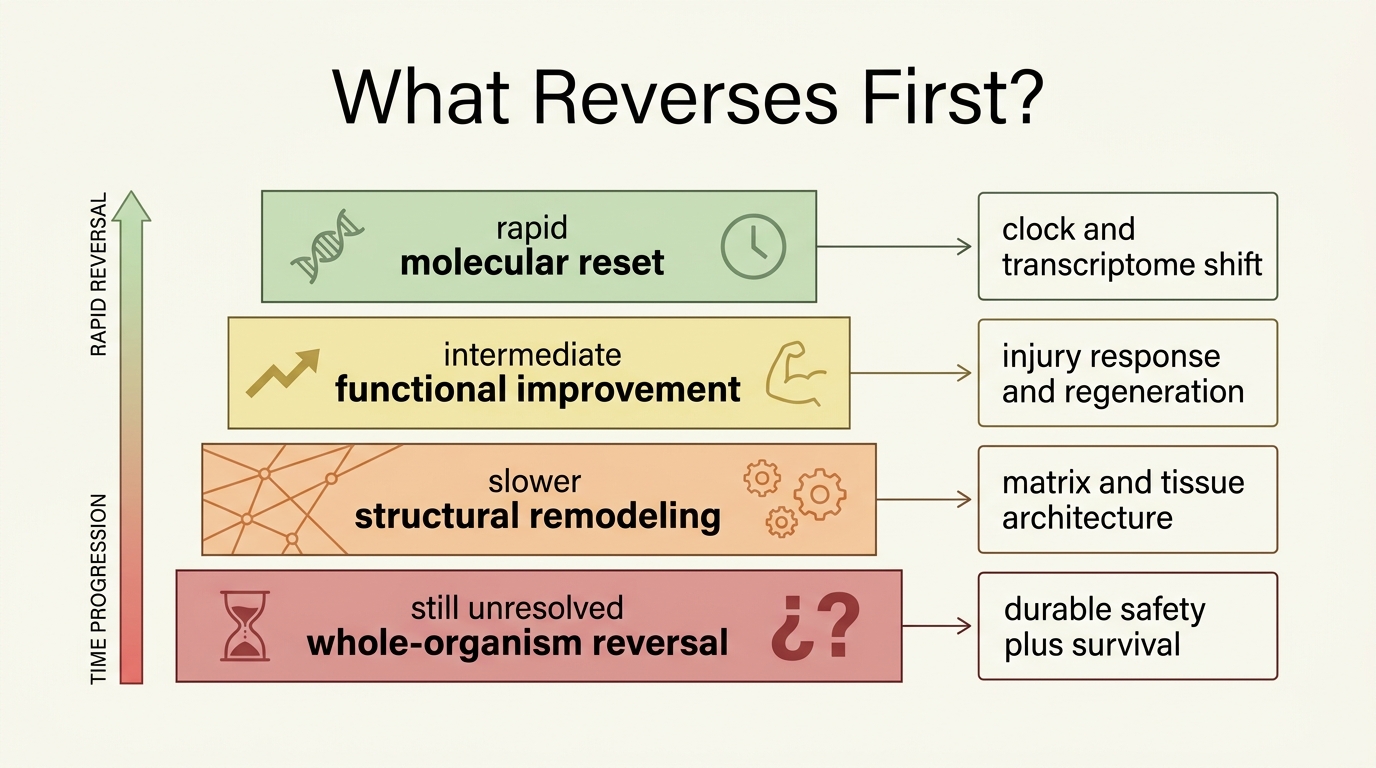
What Usually Reverses First
Animal work suggests that the first layers to move are often regulatory and signaling layers rather than gross anatomy. Epigenetic marks, transcriptional programs, inflammatory profiles, and regenerative responsiveness can shift on timescales far shorter than those required to rebuild an organ or reverse every accumulated lesion. This is exactly what one would expect if aging contains both structural damage and dynamic misregulation.
That distinction is useful because it explains both the promise and the frustration of current results. A tissue can look biologically younger by some markers while still carrying older extracellular matrix, somatic mutation burden, mitochondrial defects, clonal drift, or cancer risk. Those deeper layers constrain how far a partial reversal can propagate.
| Layer | What animal models suggest | Main limitation |
|---|---|---|
| Molecular age signatures | Often the first to move under reprogramming or systemic-environment change | Proxy movement does not guarantee durable organism-wide benefit |
| Local regenerative function | Can improve meaningfully in selected tissues and injury models | May not generalize across organs with different repair logic |
| Structural tissue remodeling | Possible but slower and less consistently demonstrated | Old scaffold, fibrosis, and mutation burden can remain |
| Whole-organism durability | Still the highest bar and least established category | Requires long-horizon safety, multi-tissue benefit, and robust survival or healthspan evidence |
Where Enthusiasm Still Runs Ahead of Evidence
The biggest overreach is treating any backward movement in a clock, transcriptome, or local function assay as if the entire aging process has been reversed. Animal studies do not justify that shortcut. They justify a narrower statement: some components of aged state remain modifiable well into late life, and some interventions can push selected tissues toward younger behavior.
A second overreach is confusing rescue after injury with generic anti-aging. The famous optic-nerve and retinal work is powerful because it demonstrates regained regenerative competence in mature neurons. But neurons under injury stress are not identical to the whole organism aging gradually across decades. The result is evidence of reversibility, not automatic proof of generalized rejuvenation.
A third overreach is collapsing short-run improvement into permanence. Many age-linked systems can be nudged transiently. The harder question is whether the organism stays younger after intervention stops, whether repeated cycling remains safe, and whether the cost of control grows as tissues age further.
What The Animal Data Probably Means
The strongest inference is that aging contains a larger reversible component than classical wear-and-tear models implied. That reversible component appears to include epigenetic state, stress-response posture, inflammatory signaling, and at least some forms of stem-cell and regenerative behavior. In other words, age is not only a pile of irreversible lesions. Part of it is a maintained operating mode.
The equally important counterpoint is that maintained operating mode is not the whole story. Old organisms still carry mutation burden, matrix changes, altered selection histories, mitochondrial bottlenecks, and tissue-specific architecture problems. That is why a real rejuvenation program will probably need layered intervention rather than one reset event. Even a successful reprogramming platform may end up as one module inside a broader repair stack.
This layered reading fits the broader LifeMeter picture. Multi-omics integration can sharpen which layers moved. Stem-cell work shows why niche quality matters alongside cell state. Matrix aging explains why youthful signaling alone may fail inside an old scaffold.
What Human Translation Would Have To Prove
To move from persuasive animal signal to serious medicine, a human program would need to prove more than clock movement. It would need durable functional gain in the relevant tissue, preserved identity, acceptable oncologic risk, controllable delivery, and a monitoring stack that can tell true rejuvenation from temporary biomarker improvement. It would also need to show that benefit persists long enough to justify intervention burden.
That is why the next decade matters more than the last one. The field no longer needs to establish that reversible signatures exist at all. It needs to establish which reversible signatures are decision-grade, which are misleading, and which can be moved safely in humans at therapeutically meaningful scale.
Bottom Line
Animal models now make one proposition hard to deny: some aspects of biological aging are reversible. The proposition they do not yet prove is that aging in its full organismal sense has become a solved engineering problem. The evidence is strongest where endpoints are narrow, tissues are tractable, and interventions are tightly controlled. That is already a major scientific shift. It is not yet a general age-reversal verdict.
Primary Source Anchors
Ocampo, A. et al. In Vivo Amelioration of Age-Associated Hallmarks by Partial Reprogramming. Cell (2016).
Lu, Y. et al. Reprogramming to recover youthful epigenetic information and restore vision. Nature (2020).
Browder, K. C. et al. In vivo partial reprogramming alters age-associated molecular changes during physiological aging in mice. Nature Aging (2022).
Heterochronic parabiosis and systemic rejuvenation literature, including recent prolonged-parabiosis molecular-aging work in mice.
Supporting context from existing LifeMeter analyses on partial reprogramming, in vivo Yamanaka-factor constraints, and methylation drift.
Translate this longevity claim into a capital-runway decision.
Life extension logic only matters if the balance sheet can carry it. Move into WealthMeter to compare assets, spending, and yield assumptions against the same long-horizon planning problem.